Diferenças entre edições de "Etileno"
| (2 edições intermédias de um utilizador não apresentadas) | |||
| Linha 1: | Linha 1: | ||
| − | <span style="font-size:8pt"><b> | + | <span style="font-size:8pt"><b>Referência : </b> Neves, M., Correia, S., Canhoto, J., (2024) ''Etileno'', [https://rce.casadasciencias.org Rev. Ciência Elem.], V12(2):013 |
| + | <br> | ||
| + | <span style="font-size:8pt"><b>Autores</b>: <i>Mariana Neves, Sandra Correia e Jorge Canhoto</i></span><br> | ||
| + | <span style="font-size:8pt"><span style="font-size:8pt"><b>Editor</b>: <i>[[Usuário:Jntavar|João Nuno Tavares]]</i></span><br> | ||
| + | <span style="font-size:8pt"><b>DOI</b>: <i>[[https://doi.org/10.24927/rce2024.013 https://doi.org/10.24927/rce2024.013]]</i></span><br> | ||
| + | <html><a href="https://rce.casadasciencias.org/rceapp/static/docs/artigos/2024-001.pdf" target="_blank"> | ||
| + | <img src="https://rce.casadasciencias.org/static/images/layout/pdf.png" alt="PDF Download"></a></html> | ||
---- | ---- | ||
| − | O etileno é uma | + | |
| + | ==Resumo== | ||
| + | |||
| + | O etileno, C<sub>2</sub>H<sub>4</sub>, é uma hormona vegetal que tem um papel crucial em vários processos do desenvolvimento das plantas e em repostas a vários fatores de stresse. O seu efeito nos tecidos vegetais é consequência de uma via de sinalização complexa que envolve a ativação de fatores de transcrição que controlam a expressão de inúmeros genes de resposta ao etileno. Esta hormona tem um papel fundamental no desenvolvimento da raiz, formação de pelos radiculares, senescência, abcisão foliar e maturação dos frutos. O etileno está também envolvido na tolerância a diversos fatores de stresses abióticos e bióticos como temperaturas elevadas, salinidade, seca e ataques de fungos e bactérias. De um modo geral, esta hormona destaca-se como um elemento-chave na regulação de processos de desenvolvimento, stresse e adaptação das plantas ao ambiente que as rodeia. | ||
| + | |||
| + | |||
| + | |||
| + | <html> | ||
| + | <p class='mainText'>O etileno é uma hormona vegetal de estrutura simples, C<sub>2</sub>H<sub>4</sub>, gasosa à temperatura ambiente, | ||
| + | que é sintetizada e libertada pelos tecidos vegetais</html><ref><html>ABELES, F. B., | ||
| + | <em>Ethylene in Plant Biology</em>, <em>New York, USA: Academic Press</em>. 1973.</html></ref><html>. Apesar da sua simplicidade, | ||
| + | participa em múltiplos processos do desenvolvimento vegetal e em várias respostas aos | ||
| + | stresses bióticos e abióticos</html><ref><html>KHAN, N. A. <em>et al.</em>, | ||
| + | <em><a class="a-link" target="_blank" | ||
| + | href="https://www.frontiersin.org/journals/plant-science/articles/10.3389/fpls.2017.01782/full">Editorial: Ethylene: A Key Regulatory Molecule in Plants</a></em>, <em>Front Plant Sci</em>, 8, 1782. 2017.</html></ref><html>. O efeito do etileno nas plantas foi reportado pela primeira | ||
| + | vez na ervilha, em 1901, por Dimitry Neljubov. No entanto, foi Richard Gane, em 1934, quem | ||
| + | fez a grande descoberta de que as plantas não só respondiam ao etileno, como também o | ||
| + | sintetizavam</html><ref><html>BAKSHI, A. <em>et al.</em>, | ||
| + | <em><a class="a-link" target="_blank" | ||
| + | href="https://www.researchgate.net/publication/279805886_History_of_Research_on_the_Plant_Hormone_Ethylene">History of Research on the Plant Hormone Ethylene</a></em>, <em>J Plant Growth Regul</em>, 34, 809-827. 2015.</html></ref><html>. A correlação entre a síntese de etileno e a sua atividade biológica mostrou à | ||
| + | comunidade científica que moléculas em estado gasoso podiam atuar também como hormonas</html><ref><html>CHANG, C., Q&A, | ||
| + | <em><a class="a-link" target="_blank" | ||
| + | href="https://bmcbiol.biomedcentral.com/articles/10.1186/s12915-016-0230-0">How do plants respond to ethylene and what is its importance?</a></em>, <em>BMC Biol</em>, 14, 7. 2016.</html></ref><html>. | ||
| + | O etileno marcou, assim, a história como a primeira molécula gasosa com função | ||
| + | sinalizadora a ser conhecida.</p> | ||
| + | |||
| + | <p class='mainText'>A biossíntese do etileno foi bastante estudada na segunda metade do século XX, período no | ||
| + | qual Adams e Yang identificaram a s-adenosilmetionina (SAM) e o ácido aminociclopropano | ||
| + | carboxílico (ACC), como os seus dois intermediários</html><ref><html>ADAMS, D. O. & YANG S. F., | ||
| + | <em><a class="a-link" target="_blank" | ||
| + | href="https://www.ncbi.nlm.nih.gov/pmc/articles/PMC382898/">Ethylene biosynthesis: Identification of 1-aminocyclopropane-1-carboxylic acid as an intermediate in the conversion of methionine to ethylene</a></em>, <em>Proc Natl Acad Sci USA</em>, 76, 170-174. 1979.</html></ref><html><sup>,</sup> </html><ref><html>ADAMS, D. O. & YANG S. F., | ||
| + | <em><a class="a-link" target="_blank" | ||
| + | href="https://pubmed.ncbi.nlm.nih.gov/16660208/">Methionine Metabolism in Apple Tissue: implication of s-adenosylmethionine as an intermediate in the conversion of methionine to ethylene</a></em>, <em>Plant Physiol</em>, 60, 892-896. 1977.</html></ref><html>. De um modo geral, a biossíntese desta | ||
| + | hormona inicia-se com a conversão de SAM em ACC, pela ACC sintase, e finalmente o ACC é | ||
| + | convertido em etileno pela ACC oxidase</html><ref><html>YANG, S. F. & HOFFMAN, N. E., | ||
| + | <em><a class="a-link" target="_blank" | ||
| + | href="https://www.annualreviews.org/content/journals/10.1146/annurev.pp.35.060184.001103">Ethylene biosynthesis and its regulation in higher plants</a></em>, <em>Annu Rev Plant Physiol</em>, 35, 155-189. 1984.</html></ref><html>. O uso de diversos mutantes com resposta variável | ||
| + | ao etileno, obtidos da planta modelo <em>Arabidopsis thaliana</em>, permitiu ao longo dos anos não só | ||
| + | compreender a função desta hormona no desenvolvimento vegetal, como também perceber | ||
| + | quais os componentes da sua via de sinalização</html><ref><html>CHEN, Y. F. <em>et al.</em>, | ||
| + | <em><a class="a-link" target="_blank" | ||
| + | href="https://academic.oup.com/aob/article/95/6/901/188924">Ethylene signal transduction</a></em>, <em>Ann Bot</em>, 95, 901-915. 2005.</html></ref><html>, dos quais se destacam os recetores de etileno. | ||
| + | O recetor de etileno denominado <em>Ethylene Receptor 1</em> (ETR1) em <em>A. thaliana</em> foi não só o | ||
| + | primeiro recetor de etileno identificado, como também foi o primeiro recetor de hormonas vegetais | ||
| + | a ser clonado e sequenciado</html><ref><html>CHANG, C. <em>et al.</em>, | ||
| + | <em><a class="a-link" target="_blank" | ||
| + | href="https://www.science.org/doi/10.1126/science.8211181">Arabidopsis Ethylene-Response Gene ETR1: Similarity of Product to Two-Component Regulators</a></em>, <em>Science</em>, 262, 539- | ||
| + | 544. 1993.</html></ref><html>. Em <em>A. thaliana</em>, o etileno é reconhecido por uma família de | ||
| + | cinco recetores denominados ETR1, ERS1 (<em>Ethylene Response Sensor 1</em>), ETR2, EIN4 (<em>Ethylene | ||
| + | Insensitive 4</em>) e ERS2</html><ref><html>BLEECKER, A. B. <em>et al.</em>, | ||
| + | <em><a class="a-link" target="_blank" | ||
| + | href="https://pubmed.ncbi.nlm.nih.gov/9800203/">The ethylene-receptor family from Arabidopsis: structure and function</a></em>, <em>Philos Trans R Soc Lond B Biol Sci</em>, 353, | ||
| + | 1405-1412. 1998.</html></ref><html>. Estes recetores situam-se na membrana do retículo endoplasmático | ||
| + | (ER) e é a partir da ligação do etileno ao recetor que se desencadeia uma via de sinalização | ||
| + | complexa</html><ref><html>BINDER, B. M., | ||
| + | <em><a class="a-link" target="_blank" | ||
| + | href="https://pubmed.ncbi.nlm.nih.gov/32332098/">Ethylene signaling in plants</a></em>, <em>J Biol.Chem</em>, 295, 7710-7725. 2020.</html></ref><html>. Dois componentes chave nesta via de sinalização são a proteína-cinase <em>Constituve | ||
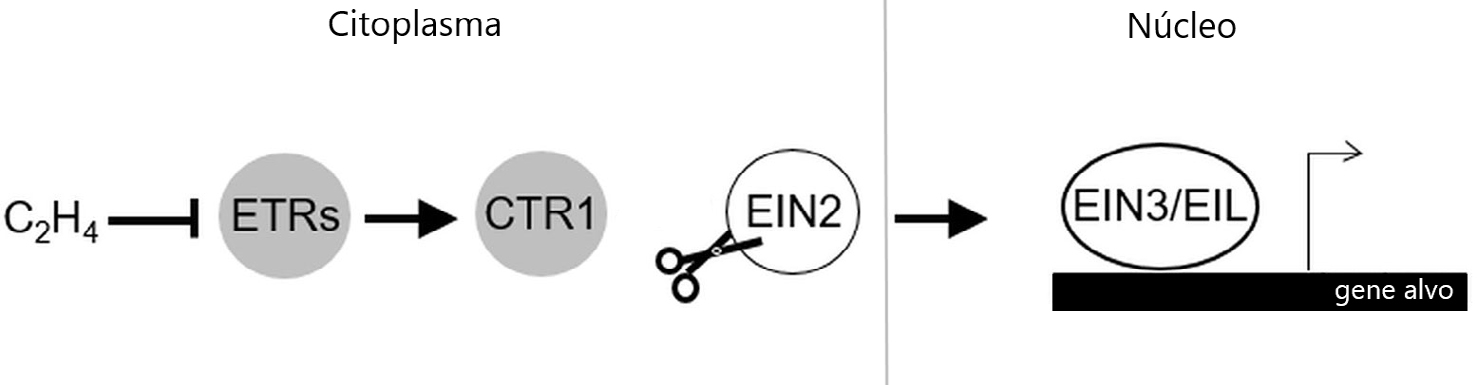
| + | Triple Response 1</em> (CTR1) e a proteína <em>Ethylene Insensitive 2</em> (EIN2), que funcionam como reguladores negativos e positivos, respetivamente (FIGURA 1). CTR1 tem a função de fosforilar | ||
| + | EIN2 e impedir a sua função na via de sinalização; já EIN2 quando não impedida tem a função | ||
| + | de aumentar a atividade de fatores de transcrição EIN3 e EIN3-LIKE 1 (EIL1) que, por sua vez, | ||
| + | controlam a transcrição de genes de resposta ao etileno, como os fatores de transcrição da | ||
| + | família ERFs (<em>Ethylene Response Factors</em>).</p> | ||
| + | |||
| + | <p class='mainText'>De uma maneira geral, na ausência de etileno (ar) os recetores mantêm a proteína-cinase | ||
| + | CTR1 ativa, que por sua vez fosforila EIN2. Esta fosforilação diminui os níveis de EIN2 impedindo | ||
| + | o seu transporte para o núcleo e a ativação dos fatores de transcrição. Na presença de | ||
| + | etileno, a sua ligação inibe os recetores, com consequente desactivação de CTR1. A perda de | ||
| + | atividade de CTR1 permite a clivagem da proteína não fosforilada EIN2, ancorada no ER. O domínio | ||
| + | C-terminal de EIN2 (EIN2-C) libertado após a clivagem, é transportado para o núcleo | ||
| + | e ativa a via de sinalização a jusante de EIN2 ao desencadear a atividade dos fatores de transcrição | ||
| + | EIN3/EIL1</html><ref><html>BINDER, B. M., | ||
| + | <em><a class="a-link" target="_blank" | ||
| + | href="https://pubmed.ncbi.nlm.nih.gov/32332098/">Ethylene signaling in plants</a></em>, <em>J Biol.Chem</em>, 295, 7710-7725. 2020.</html></ref><html><sup>,</sup> </html><ref><html>DOLGIKH, V. A. <em>et al.</em>, | ||
| + | <em><a class="a-link" target="_blank" | ||
| + | href="https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6718143/">Shaping Ethylene Response: The Role of EIN3/EIL1 Transcription Factors</a></em>, <em>Front Plant Sci</em>, 10, 1030. 2019.</html></ref><html>.</p> | ||
| + | |||
| + | <br> | ||
| + | <figure class="image-medium"> | ||
| + | <img src="https://rce.casadasciencias.org/static/images/articles/2024-013-01.jpg"> | ||
| + | </figure> | ||
| + | <center> | ||
| + | <figcaption>FIGURA 1. Esquema geral da via de sinalização do etileno e alguns dos seus componentes chave. Círculos cinzentos indicam | ||
| + | reguladores negativos; círculos brancos indicam reguladores positivos; (\(\rightarrow )) indica efeito regulador; (⊣) indica inibição. Esquema | ||
| + | adaptado de Dolgikh <em>et al.</em>, 2019</html><ref><html>DOLGIKH, V. A. <em>et al.</em>, | ||
| + | <em><a class="a-link" target="_blank" | ||
| + | href="https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6718143/">Shaping Ethylene Response: The Role of EIN3/EIL1 Transcription Factors</a></em>, <em>Front Plant Sci</em>, 10, 1030. 2019.</html></ref><html>. | ||
| + | </figcaption></center> | ||
| + | <br> | ||
| + | |||
| + | <p class='mainText'>As diferentes respostas ao etileno revelam os inúmeros papéis em que esta hormona está | ||
| + | envolvida, quer ao nível do desenvolvimento vegetal, quer nas respostas a vários fatores de | ||
| + | stress (FIGURA 2). Convém lembrar que o papel do etileno é muitas vezes resultado de uma | ||
| + | interação com outras hormonas vegetais, como as auxinas, as citocininas, as giberelinas e o | ||
| + | ácido abscísico (ABA)</html><ref><html>IQBAL, N. <em>et al.</em>, | ||
| + | <em><a class="a-link" target="_blank" | ||
| + | href="https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5378820/">Ethylene Role in Plant Growth, Development and Senescence: Interaction with Other Phytohormones</a></em>, <em>Front Plant Sci</em>, | ||
| + | 8, 475. 2017.</html></ref><html>. O efeito do etileno nas plantas começa mesmo antes da germinação | ||
| + | e tem um papel essencial no desenvolvimento da plântula</html><ref><html>AHAMMED, G. J. <em>et al.</em>, | ||
| + | <em><a class="a-link" target="_blank" | ||
| + | href="https://pubmed.ncbi.nlm.nih.gov/32220785/">Role of ethylene crosstalk in seed germination and early seedling development: A review</a></em>, <em>Plant Physiol Biochem</em>, | ||
| + | 151, 124-131. 2020.</html></ref><html>. A germinação das sementes envolve | ||
| + | inicialmente uma quebra da dormência, que resulta de um equilíbrio hormonal entre o | ||
| + | ABA e as giberelinas</html><ref><html>FINCH-SAVAGE, W. E. & LEUBNER-METZGER, G., | ||
| + | <em><a class="a-link" target="_blank" | ||
| + | href="https://pubmed.ncbi.nlm.nih.gov/16866955/">Seed dormancy and the control of germination</a></em>, <em>New Phytol</em>, 171, 501-523. 2006.</html></ref><html>. De um modo geral, o ABA mantém o fenómeno de dormência, enquanto | ||
| + | as giberelinas quebram este efeito e promovem a germinação</html><ref><html>FINCH-SAVAGE, W. E. & LEUBNER-METZGER, G., | ||
| + | <em><a class="a-link" target="_blank" | ||
| + | href="https://pubmed.ncbi.nlm.nih.gov/16866955/">Seed dormancy and the control of germination</a></em>, <em>New Phytol</em>, 171, 501-523. 2006.</html></ref><html>. O etileno participa no processo | ||
| + | de quebra da dormência, pois promove a germinação por diminuir os efeitos do ABA através | ||
| + | da regulação do seu metabolismo e via de sinalização</html><ref><html>CORBINEAU, F. <em>et al.</em>, | ||
| + | <em><a class="a-link" target="_blank" | ||
| + | href="https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4193209/">Ethylene, a key factor in the regulation of seed dormancy</a></em>, <em>Front Plant Sci</em>, 5, 539. 2014.</html></ref><html>. O etileno está também envolvido no | ||
| + | crescimento do hipocótilo, porém o seu efeito depende da presença ou ausência de luz, observando- | ||
| + | se o crescimento dos hipocótilos à luz, enquanto o seu alongamento é inibido no escuro</html><ref><html>AHAMMED, G. J. <em>et al.</em>, | ||
| + | <em><a class="a-link" target="_blank" | ||
| + | href="https://pubmed.ncbi.nlm.nih.gov/32220785/">Role of ethylene crosstalk in seed germination and early seedling development: A review</a></em>, <em>Plant Physiol Biochem</em>, | ||
| + | 151, 124-131. 2020.</html></ref><html><sup>,</sup> | ||
| + | </html><ref><html>ZHONG, S. <em>et al.</em>, | ||
| + | <em><a class="a-link" target="_blank" | ||
| + | href="https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4437768/">A molecular framework of light-controlled phytohormone action in Arabidopsis</a></em>, <em>Curr Biol</em>, 22, 1530-1535. 2012.</html></ref><html>. Ao nível do sistema radicular, o etileno tem um papel inibitório tanto no desenvolvimento | ||
| + | da raiz principal, como das raízes laterais</html><ref><html>QIN, H. & HUANG, R., | ||
| + | <em><a class="a-link" target="_blank" | ||
| + | href="https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6274790/">Auxin Controlled by Ethylene Steers Root Development</a></em>, <em>Int J Mol Sci</em>, 19, 3656. 2018.</html></ref><html>. As auxinas e o etileno inibem sinergesticamente | ||
| + | o desenvolvimento da raiz principal, no entanto, as auxinas são essenciais para a formação | ||
| + | de raízes laterais</html><ref><html>MUDAY, G. K. <em>et al.</em>, | ||
| + | <em><a class="a-link" target="_blank" | ||
| + | href="https://pubmed.ncbi.nlm.nih.gov/22406007/">Auxin and ethylene: collaborators or competitors?</a></em>, <em>Trends Plant Sci</em>, 17, 181-195. 2012.</html></ref><html>. O efeito inibitório do etileno no desenvolvimento radicular é inerente à sua | ||
| + | regulação na biossíntese, transporte e acumulação de auxinas nas diferentes zonas da raiz</html><ref><html>QIN, H. & HUANG, R., | ||
| + | <em><a class="a-link" target="_blank" | ||
| + | href="https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6274790/">Auxin Controlled by Ethylene Steers Root Development</a></em>, <em>Int J Mol Sci</em>, 19, 3656. 2018.</html></ref><html>, | ||
| + | </html><ref><html>R?ZICKA, K. <em>et al.</em>, | ||
| + | <em><a class="a-link" target="_blank" | ||
| + | href="https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1955700/">Ethylene regulates root growth through effects on auxin biosynthesis and transport-dependent auxin distribution</a></em>, <em>Plant Cell</em>, 19, 2197-2212. 2007.</html></ref><html>. Resumidamente, o etileno leva a uma acumulação de auxinas tanto na zona meristemática | ||
| + | como na zona de alongamento da raiz, resultando na inibição da proliferação e do alongamento | ||
| + | celular18. Esta acumulação de auxinas no ápice da raiz impede que estas se acumulem nas | ||
| + | zonas laterais</html><ref><html>QIN, H. & HUANG, R., | ||
| + | <em><a class="a-link" target="_blank" | ||
| + | href="https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6274790/">Auxin Controlled by Ethylene Steers Root Development</a></em>, <em>Int J Mol Sci</em>, 19, 3656. 2018.</html></ref><html><sup>,</sup> </html><ref><html>MUDAY, G. K. <em>et al.</em>, | ||
| + | <em><a class="a-link" target="_blank" | ||
| + | href="https://pubmed.ncbi.nlm.nih.gov/22406007/">Auxin and ethylene: collaborators or competitors?</a></em>, <em>Trends Plant Sci</em>, 17, 181-195. 2012.</html></ref><html>. O efeito inibitório do etileno no desenvolvimento radicular é inerente à sua | ||
| + | regulação na biossíntese, transporte e acumulação de auxinas nas diferentes zonas da raiz</html><ref><html>, onde seriam necessárias para a formação de raízes laterais. No entanto, o etileno promove a formação de pelos radiculares, pela ativação de fatores de transcrição envolvidos | ||
| + | na sua iniciação e desenvolvimento</html><ref><html>FENG, Y. <em>et al.</em>, | ||
| + | <em><a class="a-link" target="_blank" | ||
| + | href="https://pubmed.ncbi.nlm.nih.gov/29233944/">Ethylene promotes root hair growth through coordinated EIN3/EIL1 and RHD6/RSL1 activity in Arabidopsis</a></em>, <em>Proc Natl | ||
| + | Acad Sci USA</em>, 114, 13834-13839. 2017.</html></ref><html>.</p> | ||
| + | |||
| + | <p class='mainText'>O etileno é também uma hormona chave no processo de envelhecimento das plantas estando | ||
| + | envolvido no amadurecimento dos frutos e na senescência e abcisão das folhas e das | ||
| + | flores</html><ref><html>IQBAL, N. <em>et al.</em>, | ||
| + | <em><a class="a-link" target="_blank" | ||
| + | href="https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5378820/">Ethylene Role in Plant Growth, Development and Senescence: Interaction with Other Phytohormones</a></em>, <em>Front Plant Sci</em>, | ||
| + | 8, 475. 2017.</html></ref><html>. Ao nível da folha, este fenómeno está, em grande parte, relacionado com a perda de | ||
| + | clorofila</html><ref><html>GEPSTEIN, S. & THIMANN, K. V., | ||
| + | <em><a class="a-link" target="_blank" | ||
| + | href="https://www.ncbi.nlm.nih.gov/pmc/articles/PMC427489/">The Role of Ethylene in the Senescence of Oat Leaves</a></em>, <em>Plant Physiol</em>, 68, 349-354. 1981.</html></ref><html><sup>,</sup> </html><ref><html>KOUKOUNARAS, A. <em>et al.</em>, | ||
| + | <em><a class="a-link" target="_blank" | ||
| + | href="https://www.researchgate.net/publication/240417660_1-Methylcyclopropene_prevents_ethylene_induced_yellowing_of_rocket_leaves">1-Methylcyclopropene prevents ethylene induced yellowing of rocket leaves</a></em>, <em>Postharvest Biol and Technol</em>, | ||
| + | 41, 109-111. 2006.</html></ref><html>; já ao nível das flores, o etileno desencadeia um conjunto de eventos celulares nos | ||
| + | seus diferentes órgãos, que levam à sua senescência e posterior abcisão</html><ref><html>IQBAL, N. <em>et al.</em>, | ||
| + | <em><a class="a-link" target="_blank" | ||
| + | href="https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5378820/">Ethylene Role in Plant Growth, Development and Senescence: Interaction with Other Phytohormones</a></em>, <em>Front Plant Sci</em>, | ||
| + | 8, 475. 2017.</html></ref><html><sup>,</sup> </html><ref><html></html>WOLTERING, E. J. & VAN DOORN, W. G., | ||
| + | <em><a class="a-link" target="_blank" | ||
| + | href="https://oxfordjournals.org/jxb/article-abstract/39/11/1605/595917?redirectedFrom=PDF">Role of Ethylene in Senescence of Petals—Morphological and Taxonomical Relationships</a></em>, <em>J Exp | ||
| + | Bot</em>, 39, 1605–1616. 1988.</ref><html>. O amadurecimento | ||
| + | dos frutos climatéricos é o papel mais reconhecido do etileno, pois tem importantes | ||
| + | aplicações económicas</html><ref><html>KOU, X. <em>et al.</em>, | ||
| + | <em><a class="a-link" target="_blank" | ||
| + | href="https://pubmed.ncbi.nlm.nih.gov/34633626/">Different regulatory mechanisms of plant hormones in the ripening of climacteric and non-climacteric fruits: a review</a></em>, <em>Plant Mol Biol</em>, 107, 477-497. 2021.</html></ref><html>. Os frutos climatéricos são todos aqueles que apresentam uma taxa | ||
| + | de respiração elevada durante o seu amadurecimento, acompanhada de uma grande produção | ||
| + | de etileno, tais como a maçã, banana, tomate, entre ouros</html><ref><html>KOU, X. & WU, M., | ||
| + | <em><a class="a-link" target="_blank" | ||
| + | href="https://pubmed.ncbi.nlm.nih.gov/29392658/">Characterization of Climacteric and Non-Climacteric Fruit Ripening</a></em>, <em>Methods Mol Biol</em>, 1744, 89-102. 2018.</html></ref><html>. O aumento na síntese de etileno | ||
| + | e da sua perceção leva à ativação de fatores de transcrição da família ERF, que, de seguida, | ||
| + | modulam a transcrição de genes de resposta ao etileno, neste caso, genes associados ao | ||
| + | amadurecimento</html><ref><html>LIU, M. <em>et al.</em>, | ||
| + | <em><a class="a-link" target="_blank" | ||
| + | href="https://pubmed.ncbi.nlm.nih.gov/26511917/">Ethylene Control of Fruit Ripening: Revisiting the Complex Network of Transcriptional Regulation</a></em>, <em>Plant Physiol</em>, 169, 2380- | ||
| + | 2390. 2015.</html></ref><html>. Este processo leva a alteações fisiológicas e bioquímicas no fruto como a | ||
| + | redução dos níveis de clorofila e o aumento de antocianinas, modificações no metabolismo dos | ||
| + | açúcares e na biossíntese de compostos orgânicos voláteis, que resultam na mudança de cor, | ||
| + | amolecimento e aumento do aroma, sabor e conteúdo em açúcares</html><ref><html>IQBAL, N. <em>et al.</em>, | ||
| + | <em><a class="a-link" target="_blank" | ||
| + | href="https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5378820/">Ethylene Role in Plant Growth, Development and Senescence: Interaction with Other Phytohormones</a></em>, <em>Front Plant Sci</em>, | ||
| + | 8, 475. 2017.</html></ref><html><sup>,</sup> </html><ref><html>KOU, X. <em>et al.</em>, | ||
| + | <em><a class="a-link" target="_blank" | ||
| + | href="https://pubmed.ncbi.nlm.nih.gov/34633626/">Different regulatory mechanisms of plant hormones in the ripening of climacteric and non-climacteric fruits: a review</a></em>, <em>Plant Mol Biol</em>, 107, 477-497. 2021.</html></ref><html>. Controlar o processo | ||
| + | de amadurecimento dos frutos pós-colheita é fundamental para aumentar o seu tempo de | ||
| + | prateleira</html><ref><html>IQBAL, N. <em>et al.</em>, | ||
| + | <em><a class="a-link" target="_blank" | ||
| + | href="https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5378820/">Ethylene Role in Plant Growth, Development and Senescence: Interaction with Other Phytohormones</a></em>, <em>Front Plant Sci</em>, | ||
| + | 8, 475. 2017.</html></ref><html>. A estratégia mais comum passa por reduzir a síntese ou ação do etileno nos tecidos | ||
| + | vegetais; por exemplo, o composto 1-metilciclopropeno (1-MCP) tem sido usado ao longo | ||
| + | dos anos para reduzir a ação do etileno, pois atua como um inibidor competitivo para os recetores | ||
| + | de etileno</html><ref><html>BLANKENSHIP, S. M. & DOLE, J. M., | ||
| + | <em><a class="a-link" target="_blank" | ||
| + | href="https://www.researchgate.net/publication/222941993_1-Methylcyclopropene_A_review">1-Methylcyclopropene: a review</a></em>, <em>Postharvest Biol and Technol</em>, 28, 1-25. 2003.</html></ref><html>. Diminuir a temperatura e controlar a atmosfera de armazenamento a que os | ||
| + | frutos estão expostos também é uma das estratégias utilizadas, já que a redução dos níveis de | ||
| + | O<sub>2</sub> e o aumento de CO<sub>2</sub> diminui a respiração e a sensibilidade ao etileno</html><ref><html>MAHAJAN, P. V. <em>et al.</em>, | ||
| + | <em><a class="a-link" target="_blank" | ||
| + | href="https://pubmed.ncbi.nlm.nih.gov/24797137/">Postharvest treatments of fresh produce</a></em>, <em>Philos Trans A Math Phys Eng Sci</em>, 372, 20130309. 2014.</html></ref><html>.</p> | ||
| + | |||
| + | <br> | ||
| + | <figure class="image-medium"> | ||
| + | <img src="https://rce.casadasciencias.org/static/images/articles/2024-013-02.jpg"> | ||
| + | </figure><center> | ||
| + | <figcaption>FIGURA 2. À esquerda esquema de alguns papéis do etileno no desenvolvimento e tolerância a condições de stress. À direita | ||
| + | esquema de amadurecimento do tomate (baseado em Kou <em>et al.</em>. 2021</html><ref><html> KOU, X. <em>et al.</em>, | ||
| + | <em><a class="a-link" target="_blank" | ||
| + | href="https://pubmed.ncbi.nlm.nih.gov/34633626/">Different regulatory mechanisms of plant hormones in the ripening of climacteric and non-climacteric fruits: a review</a></em>, <em>Plant Mol Biol</em>, 107, 477-497. 2021.</html></ref><html>). | ||
| + | </figcaption></center> | ||
| + | <br> | ||
| + | |||
| + | <p class='mainText'>Nos últimos anos vários estudos têm reportado o papel do etileno na tolerância a vários fatores | ||
| + | de stresse, como calor</html><ref><html>POÓR, P. <em>et al.</em>, | ||
| + | <em><a class="a-link" target="_blank" | ||
| + | href="https://pubmed.ncbi.nlm.nih.gov/33713206/">Ethylene involvement in the regulation of heat stress tolerance in plants</a></em>, <em>Plant Cell Rep</em>, 41, 675-698. 2022.</html></ref><html>, hipóxia</html><ref><html>HEKHAWAT, K. <em>et al.</em>, | ||
| + | <em><a class="a-link" target="_blank" | ||
| + | href="https://www.ncbi.nlm.nih.gov/pmc/articles/PMC9818225/">Ethylene: A Master Regulator of Plant-Microbe Interactions under Abiotic Stresses</a></em>, <em>Cells</em>, 12, 31. 2022.</html></ref><html>, metais pesados</html><ref><html>KEUNEN, E. <em>et al.</em>, | ||
| + | <em><a class="a-link" target="_blank" | ||
| + | href="https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4735362/">Ethylene and Metal Stress: Small Molecule, Big Impact</a></em>, <em>Front Plant Sci</em>, 7, 23. 2016.</html></ref><html>, salinidade</html><ref><html>RIYAZUDDIN, R. <em>et al.</em>, | ||
| + | <em><a class="a-link" target="_blank" | ||
| + | href="https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7355584/">Ethylene: A Master Regulator of Salinity Stress Tolerance in Plants</a></em>, <em>Biomolecules</em>, 10, 959. 2020.</html></ref><html>, seca</html><ref><html>CUI, M. <em>et al.</em>, | ||
| + | <em><a class="a-link" target="_blank" | ||
| + | href="https://www.researchgate.net/publication/277430802_Ethylene_increases_accumulation_of_compatible_solutes_and_decreases_oxidative_stress_to_improve_plant_tolerance_to_water_stress_in_Arabidopsis">Ethylene increases accumulation of compatible solutes and decreases oxidative stress to improve plant tolerance to water stress in Arabidopsis</a></em>, <em>J Plant Biol</em>, 58, 193-201. 2015.</html></ref><html>, e ataque de bactérias, | ||
| + | fungos e herbívoros</html><ref><html>LU, J. <em>et al.</em>, | ||
| + | <em><a class="a-link" target="_blank" | ||
| + | href="https://pubmed.ncbi.nlm.nih.gov/25064847/">Contrasting effects of ethylene biosynthesis on induced plant resistance against a chewing and a piercing-sucking herbivore in rice</a></em>, <em>Mol Plant</em>, 7, 1670-1682. 2014.</html></ref><html><sup>,</sup> </html><ref><html>VAN DER ENT, S. & PIETERSE, C. M. J., | ||
| + | <em><a class="a-link" target="_blank" | ||
| + | href="https://www.researchgate.net/publication/345155987_Ethylene_Multi-Tasker_in_Plant-Attacker_Interactions">Ethylene: Multi-Tasker in Plant-Attacker Interactions</a></em>, <em>Wiley-Blackwell: New Jersey</em>, USA, 44, | ||
| + | 343-377. 2012.</html></ref><html>. No entanto, é de referir que, dependendo da espécie, o etileno | ||
| + | pode também ter uma pepel negativo nestas respostas e tornar a planta mais suscetível</html><ref><html>FATMA, M. <em>et al.</em>, | ||
| + | <em><a class="a-link" target="_blank" | ||
| + | href="https://pubmed.ncbi.nlm.nih.gov/36079592/">Ethylene Signaling under Stressful Environments: Analyzing Collaborative Knowledge</a></em>, <em>Plants</em>, 11, 2211. 2022.</html></ref><html>. A | ||
| + | tolerância a temperaturas mais elevadas promovida pelo etileno está relacionada com a homeostase | ||
| + | de espécies reativas de oxigénio (ROS), regulada por antioxidantes enzimáticos e | ||
| + | não-enzimáticos resultando em menores danos oxidativos</html><ref><html>POÓR, P. <em>et al.</em>, | ||
| + | <em><a class="a-link" target="_blank" | ||
| + | href="https://pubmed.ncbi.nlm.nih.gov/33713206/">Ethylene involvement in the regulation of heat stress tolerance in plants</a></em>, <em>Plant Cell Rep</em>, 41, 675-698. 2022.</html></ref><html>. Em condições baixas de O<sub<2</sub> o | ||
| + | etileno induz mudanças morfológicas e bioquímicas que permitem a sobrevivência da planta, | ||
| + | como a expressão de genes relacionados com o azoto e um aumento da respiração anaeróbia</html><ref><html>SHEKHAWAT, K. <em>et al.</em>, | ||
| + | <em><a class="a-link" target="_blank" | ||
| + | href="https://www.ncbi.nlm.nih.gov/pmc/articles/PMC9818225/">Ethylene: A Master Regulator of Plant-Microbe Interactions under Abiotic Stresses</a></em>, <em>Cells</em>, 12, 31. 2022.</html></ref><html>. | ||
| + | A tolerância a metais pesados, como o arsénio</html><ref><html>FU, S. F. <em>et al.</em>, | ||
| + | <em><a class="a-link" target="_blank" | ||
| + | href="https://pubmed.ncbi.nlm.nih.gov/24734953/">Transcriptome profiling of genes and pathways associated with arsenic toxicity and tolerance in Arabidopsis</a></em>, <em>BMC Plant | ||
| + | Biol</em>, 14, 94. 2014.</html></ref><html> e o cádmio</html><ref><html>CAO, F. <em>et al.</em>, | ||
| + | <em><a class="a-link" target="_blank" | ||
| + | href="https://pubmed.ncbi.nlm.nih.gov/25038590/">Genome-wide transcriptome and functional analysis of two contrasting genotypes reveals key genes for cadmium tolerance in barley</a></em>, <em>BMC Biol</em>, 15, 611. 2014.</html></ref><html><sup>,</sup> </html><ref><html>GUAN, C. <em>et al.</em>, | ||
| + | <em><a class="a-link" target="_blank" | ||
| + | href="https://www.semanticscholar.org/paper/The-glutathione-synthesis-may-be-regulated-by-in-of-Guan-Ji/2fded93dd9ca3c3c7fff77677ca79ebf87c59606">The glutathione synthesis may be regulated by cadmium-induced endogenous ethylene in Lycium chinense, and overexpression of an ethylene responsive transcription factor gene enhances tolerance to cadmium stress in tobacco</a></em>, <em>Molecular Breeding</em>, | ||
| + | 35, 123. 2015.</html></ref><html>, parece estar relacionada com | ||
| + | a modulação da transcrição de ERFs específicos resultantes da ativação da via de sinalização | ||
| + | do etileno. O etileno promove a tolerância à salinidade através da manutenção da homeostase | ||
| + | de Na<sup>+</sup>/K<sup>+</sup>, nutrientes e ROS, e também pelo aumento da assimilação de nitratos e sulfatos. A | ||
| + | tolerância à seca parece estar associada a uma regulação positiva no aumento do conteúdo em | ||
| + | açúcares e prolina de modo a combater o stress osmótico, levando a uma diminuição da peroxidação | ||
| + | lípica</html><ref><html>CUI, M. <em>et al.</em>, | ||
| + | <em><a class="a-link" target="_blank" | ||
| + | href="https://www.researchgate.net/publication/277430802_Ethylene_increases_accumulation_of_compatible_solutes_and_decreases_oxidative_stress_to_improve_plant_tolerance_to_water_stress_in_Arabidopsis">Ethylene increases accumulation of compatible solutes and decreases oxidative stress to improve plant tolerance to water stress in Arabidopsis</a></em>, <em>J Plant Biol</em>, 58, 193-201. 2015.</html></ref><html>. O ataque de agentes patogénicos, como bactérias e fungos é acompanhado | ||
| + | pela ativação de fatores de transcrição que estão envolvidos na regulação de genes associados | ||
| + | à imunidade, tornando a planta menos suscetível31.</p> | ||
| + | |||
| + | <p class='mainText'>Numa altura em que as preocupações relacionadas às alterações climáticas e à segurança | ||
| + | alimentar estão em destaque, o etileno revela-se como uma hormona chave na regulação da | ||
| + | capacidade adaptativa das plantas. Assim, é essencial compreender os múltiplos papéis do | ||
| + | etileno e os seus mecanismos de sinalização para encontrar novas estratégias que otimizem o | ||
| + | desenvolvimento das plantas e a sua adaptação a ambientes pouco favoráveis.</p> | ||
| + | |||
| + | <p> </p> | ||
| + | </html> | ||
| + | |||
| + | =Referências= | ||
| + | <references/> | ||
<br> Palavras chave: fitohormonas, ABA, giberelina, citocinina, auxina | <br> Palavras chave: fitohormonas, ABA, giberelina, citocinina, auxina | ||
| Linha 10: | Linha 289: | ||
Materiais relacionados disponíveis na [http://www.casadasciencias.org Casa das Ciências]:<br> | Materiais relacionados disponíveis na [http://www.casadasciencias.org Casa das Ciências]:<br> | ||
# [http://www.casadasciencias.org/index.php?option=com_docman&task=doc_details&gid=34734662&Itemid=23 Ação do Etileno na maturação dos frutos], porque é que os frutos ficam mais maduros se deixados ao ar? | # [http://www.casadasciencias.org/index.php?option=com_docman&task=doc_details&gid=34734662&Itemid=23 Ação do Etileno na maturação dos frutos], porque é que os frutos ficam mais maduros se deixados ao ar? | ||
| − | ---- <br>Criada em | + | <br>---- <br>Criada em 11 de Setembro de 2023<br> Revista em 7 de Dezembro de 2023<br>Aceite pelo editor em 29 de Julho de 2024<br><br> |
Edição actual desde as 13h00min de 26 de agosto de 2024
Referência : Neves, M., Correia, S., Canhoto, J., (2024) Etileno, Rev. Ciência Elem., V12(2):013
Autores: Mariana Neves, Sandra Correia e Jorge Canhoto
Editor: João Nuno Tavares
DOI: [https://doi.org/10.24927/rce2024.013]

Resumo
O etileno, C2H4, é uma hormona vegetal que tem um papel crucial em vários processos do desenvolvimento das plantas e em repostas a vários fatores de stresse. O seu efeito nos tecidos vegetais é consequência de uma via de sinalização complexa que envolve a ativação de fatores de transcrição que controlam a expressão de inúmeros genes de resposta ao etileno. Esta hormona tem um papel fundamental no desenvolvimento da raiz, formação de pelos radiculares, senescência, abcisão foliar e maturação dos frutos. O etileno está também envolvido na tolerância a diversos fatores de stresses abióticos e bióticos como temperaturas elevadas, salinidade, seca e ataques de fungos e bactérias. De um modo geral, esta hormona destaca-se como um elemento-chave na regulação de processos de desenvolvimento, stresse e adaptação das plantas ao ambiente que as rodeia.
O etileno é uma hormona vegetal de estrutura simples, C2H4, gasosa à temperatura ambiente, que é sintetizada e libertada pelos tecidos vegetais[1]. Apesar da sua simplicidade, participa em múltiplos processos do desenvolvimento vegetal e em várias respostas aos stresses bióticos e abióticos[2]. O efeito do etileno nas plantas foi reportado pela primeira vez na ervilha, em 1901, por Dimitry Neljubov. No entanto, foi Richard Gane, em 1934, quem fez a grande descoberta de que as plantas não só respondiam ao etileno, como também o sintetizavam[3]. A correlação entre a síntese de etileno e a sua atividade biológica mostrou à comunidade científica que moléculas em estado gasoso podiam atuar também como hormonas[4]. O etileno marcou, assim, a história como a primeira molécula gasosa com função sinalizadora a ser conhecida.
A biossíntese do etileno foi bastante estudada na segunda metade do século XX, período no qual Adams e Yang identificaram a s-adenosilmetionina (SAM) e o ácido aminociclopropano carboxílico (ACC), como os seus dois intermediários[5], [6]. De um modo geral, a biossíntese desta hormona inicia-se com a conversão de SAM em ACC, pela ACC sintase, e finalmente o ACC é convertido em etileno pela ACC oxidase[7]. O uso de diversos mutantes com resposta variável ao etileno, obtidos da planta modelo Arabidopsis thaliana, permitiu ao longo dos anos não só compreender a função desta hormona no desenvolvimento vegetal, como também perceber quais os componentes da sua via de sinalização[8], dos quais se destacam os recetores de etileno. O recetor de etileno denominado Ethylene Receptor 1 (ETR1) em A. thaliana foi não só o primeiro recetor de etileno identificado, como também foi o primeiro recetor de hormonas vegetais a ser clonado e sequenciado[9]. Em A. thaliana, o etileno é reconhecido por uma família de cinco recetores denominados ETR1, ERS1 (Ethylene Response Sensor 1), ETR2, EIN4 (Ethylene Insensitive 4) e ERS2[10]. Estes recetores situam-se na membrana do retículo endoplasmático (ER) e é a partir da ligação do etileno ao recetor que se desencadeia uma via de sinalização complexa[11]. Dois componentes chave nesta via de sinalização são a proteína-cinase Constituve Triple Response 1 (CTR1) e a proteína Ethylene Insensitive 2 (EIN2), que funcionam como reguladores negativos e positivos, respetivamente (FIGURA 1). CTR1 tem a função de fosforilar EIN2 e impedir a sua função na via de sinalização; já EIN2 quando não impedida tem a função de aumentar a atividade de fatores de transcrição EIN3 e EIN3-LIKE 1 (EIL1) que, por sua vez, controlam a transcrição de genes de resposta ao etileno, como os fatores de transcrição da família ERFs (Ethylene Response Factors).
De uma maneira geral, na ausência de etileno (ar) os recetores mantêm a proteína-cinase CTR1 ativa, que por sua vez fosforila EIN2. Esta fosforilação diminui os níveis de EIN2 impedindo o seu transporte para o núcleo e a ativação dos fatores de transcrição. Na presença de etileno, a sua ligação inibe os recetores, com consequente desactivação de CTR1. A perda de atividade de CTR1 permite a clivagem da proteína não fosforilada EIN2, ancorada no ER. O domínio C-terminal de EIN2 (EIN2-C) libertado após a clivagem, é transportado para o núcleo e ativa a via de sinalização a jusante de EIN2 ao desencadear a atividade dos fatores de transcrição EIN3/EIL1[12], [13].
As diferentes respostas ao etileno revelam os inúmeros papéis em que esta hormona está
envolvida, quer ao nível do desenvolvimento vegetal, quer nas respostas a vários fatores de
stress (FIGURA 2). Convém lembrar que o papel do etileno é muitas vezes resultado de uma
interação com outras hormonas vegetais, como as auxinas, as citocininas, as giberelinas e o
ácido abscísico (ABA)[15]. O efeito do etileno nas plantas começa mesmo antes da germinação
e tem um papel essencial no desenvolvimento da plântula[16]. A germinação das sementes envolve
inicialmente uma quebra da dormência, que resulta de um equilíbrio hormonal entre o
ABA e as giberelinas[17]. De um modo geral, o ABA mantém o fenómeno de dormência, enquanto
as giberelinas quebram este efeito e promovem a germinação[18]. O etileno participa no processo
de quebra da dormência, pois promove a germinação por diminuir os efeitos do ABA através
da regulação do seu metabolismo e via de sinalização[19]. O etileno está também envolvido no
crescimento do hipocótilo, porém o seu efeito depende da presença ou ausência de luz, observando-
se o crescimento dos hipocótilos à luz, enquanto o seu alongamento é inibido no escuro[20],
[21]. Ao nível do sistema radicular, o etileno tem um papel inibitório tanto no desenvolvimento
da raiz principal, como das raízes laterais[22]. As auxinas e o etileno inibem sinergesticamente
o desenvolvimento da raiz principal, no entanto, as auxinas são essenciais para a formação
de raízes laterais[23]. O efeito inibitório do etileno no desenvolvimento radicular é inerente à sua
regulação na biossíntese, transporte e acumulação de auxinas nas diferentes zonas da raiz[24],
[25]. Resumidamente, o etileno leva a uma acumulação de auxinas tanto na zona meristemática
como na zona de alongamento da raiz, resultando na inibição da proliferação e do alongamento
celular18. Esta acumulação de auxinas no ápice da raiz impede que estas se acumulem nas
zonas laterais[26], [27]. O efeito inibitório do etileno no desenvolvimento radicular é inerente à sua
regulação na biossíntese, transporte e acumulação de auxinas nas diferentes zonas da raizErro de citação: </ref> de fecho em falta, para o elemento <ref>.
O etileno é também uma hormona chave no processo de envelhecimento das plantas estando envolvido no amadurecimento dos frutos e na senescência e abcisão das folhas e das flores[28]. Ao nível da folha, este fenómeno está, em grande parte, relacionado com a perda de clorofila[29], [30]; já ao nível das flores, o etileno desencadeia um conjunto de eventos celulares nos seus diferentes órgãos, que levam à sua senescência e posterior abcisão[31], [32]. O amadurecimento dos frutos climatéricos é o papel mais reconhecido do etileno, pois tem importantes aplicações económicas[33]. Os frutos climatéricos são todos aqueles que apresentam uma taxa de respiração elevada durante o seu amadurecimento, acompanhada de uma grande produção de etileno, tais como a maçã, banana, tomate, entre ouros[34]. O aumento na síntese de etileno e da sua perceção leva à ativação de fatores de transcrição da família ERF, que, de seguida, modulam a transcrição de genes de resposta ao etileno, neste caso, genes associados ao amadurecimento[35]. Este processo leva a alteações fisiológicas e bioquímicas no fruto como a redução dos níveis de clorofila e o aumento de antocianinas, modificações no metabolismo dos açúcares e na biossíntese de compostos orgânicos voláteis, que resultam na mudança de cor, amolecimento e aumento do aroma, sabor e conteúdo em açúcares[36], [37]. Controlar o processo de amadurecimento dos frutos pós-colheita é fundamental para aumentar o seu tempo de prateleira[38]. A estratégia mais comum passa por reduzir a síntese ou ação do etileno nos tecidos vegetais; por exemplo, o composto 1-metilciclopropeno (1-MCP) tem sido usado ao longo dos anos para reduzir a ação do etileno, pois atua como um inibidor competitivo para os recetores de etileno[39]. Diminuir a temperatura e controlar a atmosfera de armazenamento a que os frutos estão expostos também é uma das estratégias utilizadas, já que a redução dos níveis de O2 e o aumento de CO2 diminui a respiração e a sensibilidade ao etileno[40].
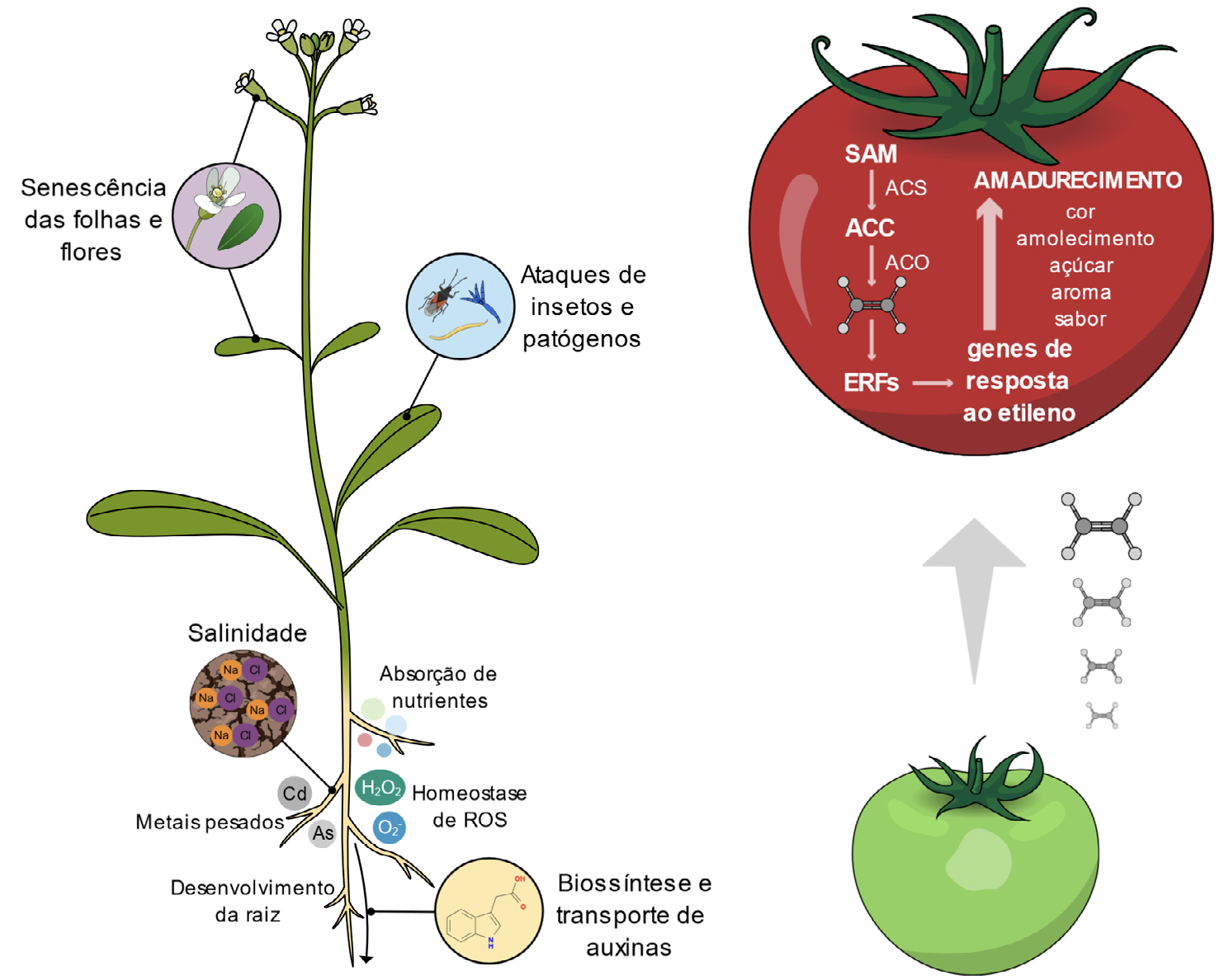
Nos últimos anos vários estudos têm reportado o papel do etileno na tolerância a vários fatores
de stresse, como calor[42], hipóxia[43], metais pesados[44], salinidade[45], seca[46], e ataque de bactérias,
fungos e herbívoros[47], [48]. No entanto, é de referir que, dependendo da espécie, o etileno
pode também ter uma pepel negativo nestas respostas e tornar a planta mais suscetível[49]. A
tolerância a temperaturas mais elevadas promovida pelo etileno está relacionada com a homeostase
de espécies reativas de oxigénio (ROS), regulada por antioxidantes enzimáticos e
não-enzimáticos resultando em menores danos oxidativos[50]. Em condições baixas de O
Numa altura em que as preocupações relacionadas às alterações climáticas e à segurança alimentar estão em destaque, o etileno revela-se como uma hormona chave na regulação da capacidade adaptativa das plantas. Assim, é essencial compreender os múltiplos papéis do etileno e os seus mecanismos de sinalização para encontrar novas estratégias que otimizem o desenvolvimento das plantas e a sua adaptação a ambientes pouco favoráveis.
Referências
- ↑ ABELES, F. B., Ethylene in Plant Biology, New York, USA: Academic Press. 1973.
- ↑ KHAN, N. A. et al., Editorial: Ethylene: A Key Regulatory Molecule in Plants, Front Plant Sci, 8, 1782. 2017.
- ↑ BAKSHI, A. et al., History of Research on the Plant Hormone Ethylene, J Plant Growth Regul, 34, 809-827. 2015.
- ↑ CHANG, C., Q&A, How do plants respond to ethylene and what is its importance?, BMC Biol, 14, 7. 2016.
- ↑ ADAMS, D. O. & YANG S. F., Ethylene biosynthesis: Identification of 1-aminocyclopropane-1-carboxylic acid as an intermediate in the conversion of methionine to ethylene, Proc Natl Acad Sci USA, 76, 170-174. 1979.
- ↑ ADAMS, D. O. & YANG S. F., Methionine Metabolism in Apple Tissue: implication of s-adenosylmethionine as an intermediate in the conversion of methionine to ethylene, Plant Physiol, 60, 892-896. 1977.
- ↑ YANG, S. F. & HOFFMAN, N. E., Ethylene biosynthesis and its regulation in higher plants, Annu Rev Plant Physiol, 35, 155-189. 1984.
- ↑ CHEN, Y. F. et al., Ethylene signal transduction, Ann Bot, 95, 901-915. 2005.
- ↑ CHANG, C. et al., Arabidopsis Ethylene-Response Gene ETR1: Similarity of Product to Two-Component Regulators, Science, 262, 539- 544. 1993.
- ↑ BLEECKER, A. B. et al., The ethylene-receptor family from Arabidopsis: structure and function, Philos Trans R Soc Lond B Biol Sci, 353, 1405-1412. 1998.
- ↑ BINDER, B. M., Ethylene signaling in plants, J Biol.Chem, 295, 7710-7725. 2020.
- ↑ BINDER, B. M., Ethylene signaling in plants, J Biol.Chem, 295, 7710-7725. 2020.
- ↑ DOLGIKH, V. A. et al., Shaping Ethylene Response: The Role of EIN3/EIL1 Transcription Factors, Front Plant Sci, 10, 1030. 2019.
- ↑ DOLGIKH, V. A. et al., Shaping Ethylene Response: The Role of EIN3/EIL1 Transcription Factors, Front Plant Sci, 10, 1030. 2019.
- ↑ IQBAL, N. et al., Ethylene Role in Plant Growth, Development and Senescence: Interaction with Other Phytohormones, Front Plant Sci, 8, 475. 2017.
- ↑ AHAMMED, G. J. et al., Role of ethylene crosstalk in seed germination and early seedling development: A review, Plant Physiol Biochem, 151, 124-131. 2020.
- ↑ FINCH-SAVAGE, W. E. & LEUBNER-METZGER, G., Seed dormancy and the control of germination, New Phytol, 171, 501-523. 2006.
- ↑ FINCH-SAVAGE, W. E. & LEUBNER-METZGER, G., Seed dormancy and the control of germination, New Phytol, 171, 501-523. 2006.
- ↑ CORBINEAU, F. et al., Ethylene, a key factor in the regulation of seed dormancy, Front Plant Sci, 5, 539. 2014.
- ↑ AHAMMED, G. J. et al., Role of ethylene crosstalk in seed germination and early seedling development: A review, Plant Physiol Biochem, 151, 124-131. 2020.
- ↑ ZHONG, S. et al., A molecular framework of light-controlled phytohormone action in Arabidopsis, Curr Biol, 22, 1530-1535. 2012.
- ↑ QIN, H. & HUANG, R., Auxin Controlled by Ethylene Steers Root Development, Int J Mol Sci, 19, 3656. 2018.
- ↑ MUDAY, G. K. et al., Auxin and ethylene: collaborators or competitors?, Trends Plant Sci, 17, 181-195. 2012.
- ↑ QIN, H. & HUANG, R., Auxin Controlled by Ethylene Steers Root Development, Int J Mol Sci, 19, 3656. 2018.
- ↑ R?ZICKA, K. et al., Ethylene regulates root growth through effects on auxin biosynthesis and transport-dependent auxin distribution, Plant Cell, 19, 2197-2212. 2007.
- ↑ QIN, H. & HUANG, R., Auxin Controlled by Ethylene Steers Root Development, Int J Mol Sci, 19, 3656. 2018.
- ↑ MUDAY, G. K. et al., Auxin and ethylene: collaborators or competitors?, Trends Plant Sci, 17, 181-195. 2012.
- ↑ IQBAL, N. et al., Ethylene Role in Plant Growth, Development and Senescence: Interaction with Other Phytohormones, Front Plant Sci, 8, 475. 2017.
- ↑ GEPSTEIN, S. & THIMANN, K. V., The Role of Ethylene in the Senescence of Oat Leaves, Plant Physiol, 68, 349-354. 1981.
- ↑ KOUKOUNARAS, A. et al., 1-Methylcyclopropene prevents ethylene induced yellowing of rocket leaves, Postharvest Biol and Technol, 41, 109-111. 2006.
- ↑ IQBAL, N. et al., Ethylene Role in Plant Growth, Development and Senescence: Interaction with Other Phytohormones, Front Plant Sci, 8, 475. 2017.
- ↑ WOLTERING, E. J. & VAN DOORN, W. G., <a class="a-link" target="_blank" href="https://oxfordjournals.org/jxb/article-abstract/39/11/1605/595917?redirectedFrom=PDF">Role of Ethylene in Senescence of Petals—Morphological and Taxonomical Relationships</a>, J Exp Bot, 39, 1605–1616. 1988.
- ↑ KOU, X. et al., Different regulatory mechanisms of plant hormones in the ripening of climacteric and non-climacteric fruits: a review, Plant Mol Biol, 107, 477-497. 2021.
- ↑ KOU, X. & WU, M., Characterization of Climacteric and Non-Climacteric Fruit Ripening, Methods Mol Biol, 1744, 89-102. 2018.
- ↑ LIU, M. et al., Ethylene Control of Fruit Ripening: Revisiting the Complex Network of Transcriptional Regulation, Plant Physiol, 169, 2380- 2390. 2015.
- ↑ IQBAL, N. et al., Ethylene Role in Plant Growth, Development and Senescence: Interaction with Other Phytohormones, Front Plant Sci, 8, 475. 2017.
- ↑ KOU, X. et al., Different regulatory mechanisms of plant hormones in the ripening of climacteric and non-climacteric fruits: a review, Plant Mol Biol, 107, 477-497. 2021.
- ↑ IQBAL, N. et al., Ethylene Role in Plant Growth, Development and Senescence: Interaction with Other Phytohormones, Front Plant Sci, 8, 475. 2017.
- ↑ BLANKENSHIP, S. M. & DOLE, J. M., 1-Methylcyclopropene: a review, Postharvest Biol and Technol, 28, 1-25. 2003.
- ↑ MAHAJAN, P. V. et al., Postharvest treatments of fresh produce, Philos Trans A Math Phys Eng Sci, 372, 20130309. 2014.
- ↑ KOU, X. et al., Different regulatory mechanisms of plant hormones in the ripening of climacteric and non-climacteric fruits: a review, Plant Mol Biol, 107, 477-497. 2021.
- ↑ POÓR, P. et al., Ethylene involvement in the regulation of heat stress tolerance in plants, Plant Cell Rep, 41, 675-698. 2022.
- ↑ HEKHAWAT, K. et al., Ethylene: A Master Regulator of Plant-Microbe Interactions under Abiotic Stresses, Cells, 12, 31. 2022.
- ↑ KEUNEN, E. et al., Ethylene and Metal Stress: Small Molecule, Big Impact, Front Plant Sci, 7, 23. 2016.
- ↑ RIYAZUDDIN, R. et al., Ethylene: A Master Regulator of Salinity Stress Tolerance in Plants, Biomolecules, 10, 959. 2020.
- ↑ CUI, M. et al., Ethylene increases accumulation of compatible solutes and decreases oxidative stress to improve plant tolerance to water stress in Arabidopsis, J Plant Biol, 58, 193-201. 2015.
- ↑ LU, J. et al., Contrasting effects of ethylene biosynthesis on induced plant resistance against a chewing and a piercing-sucking herbivore in rice, Mol Plant, 7, 1670-1682. 2014.
- ↑ VAN DER ENT, S. & PIETERSE, C. M. J., Ethylene: Multi-Tasker in Plant-Attacker Interactions, Wiley-Blackwell: New Jersey, USA, 44, 343-377. 2012.
- ↑ FATMA, M. et al., Ethylene Signaling under Stressful Environments: Analyzing Collaborative Knowledge, Plants, 11, 2211. 2022.
- ↑ POÓR, P. et al., Ethylene involvement in the regulation of heat stress tolerance in plants, Plant Cell Rep, 41, 675-698. 2022.
- ↑ SHEKHAWAT, K. et al., Ethylene: A Master Regulator of Plant-Microbe Interactions under Abiotic Stresses, Cells, 12, 31. 2022.
- ↑ FU, S. F. et al., Transcriptome profiling of genes and pathways associated with arsenic toxicity and tolerance in Arabidopsis, BMC Plant Biol, 14, 94. 2014.
- ↑ CAO, F. et al., Genome-wide transcriptome and functional analysis of two contrasting genotypes reveals key genes for cadmium tolerance in barley, BMC Biol, 15, 611. 2014.
- ↑ GUAN, C. et al., The glutathione synthesis may be regulated by cadmium-induced endogenous ethylene in Lycium chinense, and overexpression of an ethylene responsive transcription factor gene enhances tolerance to cadmium stress in tobacco, Molecular Breeding, 35, 123. 2015.
- ↑ CUI, M. et al., Ethylene increases accumulation of compatible solutes and decreases oxidative stress to improve plant tolerance to water stress in Arabidopsis, J Plant Biol, 58, 193-201. 2015.
Palavras chave: fitohormonas, ABA, giberelina, citocinina, auxina
Materiais relacionados disponíveis na Casa das Ciências:
- Ação do Etileno na maturação dos frutos, porque é que os frutos ficam mais maduros se deixados ao ar?
----
Criada em 11 de Setembro de 2023
Revista em 7 de Dezembro de 2023
Aceite pelo editor em 29 de Julho de 2024
