Diversidade no reino Epigenética, Darwin e Lamarck
Referência : Arcanjo, F. G., Silva, E. P., (2018) Diversidade no reino Epigenética, Darwin e Lamarck, Rev. Ciência Elem., V6(3):067
Autores: Fernanda Gonçalves Arcanjo e Edson Pereira da Silva
Editor: José Ferreira Gomes
DOI: [http://doi.org/10.24927/rce2018.067]

O termo “epigenética” diz respeito aos mecanismos através dos quais uma célula adquire sua forma e/ou função e como essa forma e função se mantêm estáveis numa linhagem de células. Portanto, a epigenética estuda todas as interações moleculares que definem a diferenciação celular, tanto aquelas de nível nuclear (associado ao DNA) e citoplasmático quanto as interações célula-célula e dessas com o ambiente. Dentre as linhas de pesquisa em epigenética, a herança epigenética transgeracional vem ganhando bastante destaque na atualidade. Este fenômeno se refere às alterações nos padrões de expressão dos genes que são herdadas através das gerações sem que sejam determinadas por mudanças na sequência de bases do DNA. Por isso, estes estudos vêm sendo apelidados de uma “volta ao Lamarckismo”. Neste artigo, vamos tentar explicitar melhor do que se trata a epigenética e questionar sua frequente associação ao nome de Lamarck frente ao paradigma darwiniano na Biologia.
De origem grega, o prefixo epi significa “sobre”, portanto, a palavra epigenética pode ser traduzida como àquilo que está sobre/em cima/além do gene ou, mais especificamente, da molécula de DNA. No entanto, desde a sua proposição pelo geneticista Conrad Waddington, em 1942, o termo “epigenética” foi utilizado em diferentes sentidos. Inicialmente, quando a epigenética ainda não se configurava como um campo de estudo, havia, apenas, um interesse de pesquisadores nas áreas da genética e da biologia do desenvolvimento nos fatores responsáveis por promover a diferenciação celular a partir de um único genoma. Eles queriam entender como que um mesmo indivíduo poderia ter uma quantidade tão variada de tipos celulares se a informação genética de todas as células é a mesma.
Naquele momento buscavam-se explicações apenas ao nível mitótico de hereditariedade. Ou seja, em meados do século XX, estes pesquisadores estavam preocupados somente em compreender como as características epigenéticas eram passadas da célula-mãe para as células-filhas, permitindo a constituição dos diferentes tecidos e órgãos. Ainda não se falava muito sobre como isso era passado através das gerações, pois tal questão ainda era um problema do desenvolvimento do organismo e não da genética. Por isso mesmo, a associação destes estudos com o termo epigenética era rara[1].
Posteriormente, a epigenética adquiriu uma definição mais elaborada e passou a disputar espaço teórico com a genética do desenvolvimento, uma disciplina que, ao contrário da epigenética, estava preocupada com o acoplamento da variação genotípica (variação a nível molecular, presente na molécula de DNA) à variação fenotípica (variação visível). Assim sendo, de um lado estava a biologia do desenvolvimento, que queria entender como as características que eram expressas fisicamente estavam codificadas no DNA e, do outro lado, estava a epigenética, preocupada em entender os fenômenos pelos quais a variação no genótipo não produzia nenhuma variação sobre o fenótipo, ou quando a variação do fenótipo não poderia ser explicada por uma variação no DNA[2].
Hoje, os estudos voltados para o desenvolvimento envolvendo expressão gênica já pressupõem a existência de redes complexas de interação entre genes e fatores moleculares epigenéticos dando origem às variações fenotípicas, superando esta contradição ingênua, do final do século XX, entre as perspectivas genética e epigenética do desenvolvimento.
Os dois principais e mais estudados mecanismos de marcação epigenética sobre o DNA são a metilação (associação de um radical metil a uma região específica do genoma) e a condensação do cromossomo (processo pelo qual a molécula de DNA se enrola sobre ela mesma junto à um conjunto proteico, diminuindo seu tamanho e área de contato) e ambos fenômenos estão associados, especialmente em mamíferos, à inibição da expressão de diversos genes[3]. Porém, estes são apenas dois exemplos de fenômenos epigenéticos em sua definição mais simples.
Há, ainda, o problema das origens das marcações epigenéticas e discute-se muito sobre a influência dos fatores ambientais (tudo que é externo ao DNA, desde as condições do citoplasma até os fatores climáticos etc.) neste processo[4]. Um experimento clássico discutindo esta questão da epigenética é o de Waterland e Jirtle[5] com ratos agouti (FIGURA 1). Neste sentido, embora o campo de estudo da epigenética se encontre, atualmente, bem estabelecido, a palavra ainda é utilizada em sentidos diversos, bem diferentes daquele cunhado por Waddington, devido a variedade e complexidade dos fenômenos que são entendidos, hoje, como epigenéticos. E, entre todas as novidades evidenciadas pela epigenética, uma vem se tornando particularmente popular e controversa: o fenômeno da herança epigenética (ou herança meiótica ou, ainda, herança epigenética transgeracional).
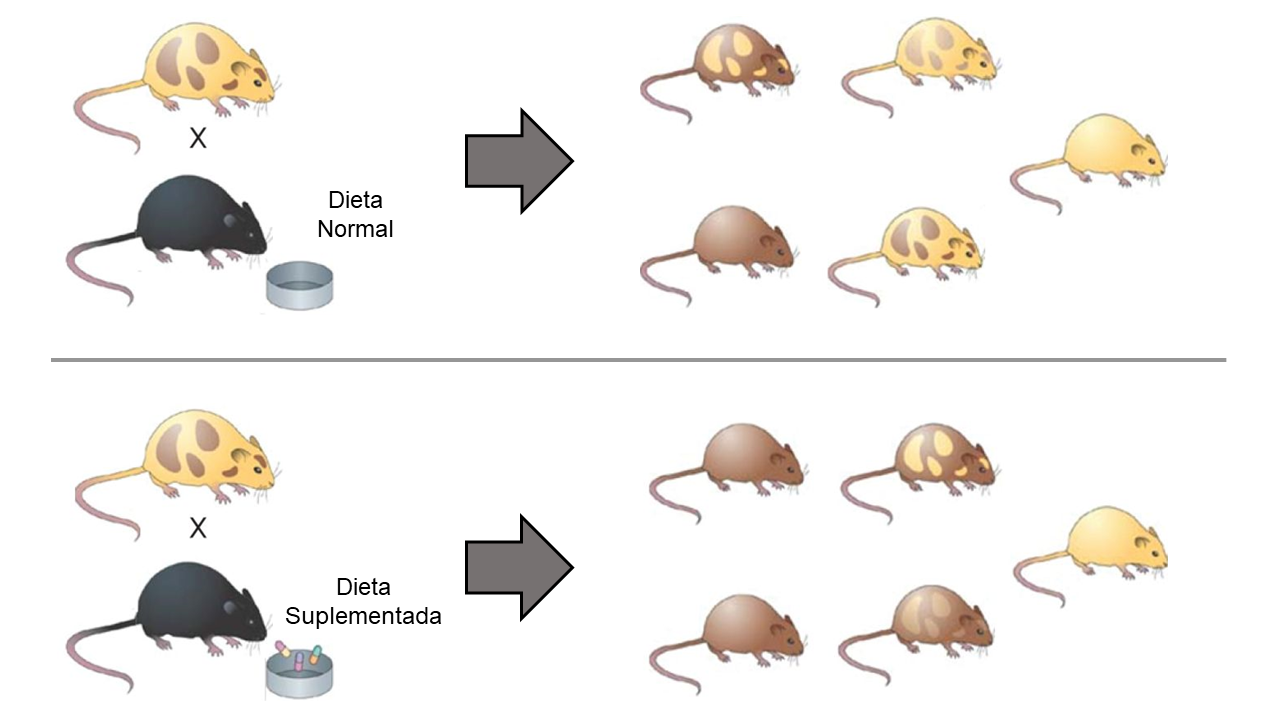
A herança epigenética é, justamente, o fenômeno de herança de marcações epigenéticas, não somente no nível do desenvolvimento (nível celular, mitótico), mas através de gerações. O que se sabe atualmente sobre este fenômeno é que nem toda a variação epigenética é herdada e, muitas vezes, quando herdada, se mantém por poucas gerações. Porém, já foram evidenciados casos de herança epigenética estável por dezenas de gerações, o que promove hoje intensa discussão a respeito da influência que esse fenômeno pode ter sobre o processo evolutivo[6], sendo especialmente polêmica a questão do papel do ambiente na produção de marcações epigenéticas herdáveis.
Curiosamente, herança epigenética é para muitos, hoje, sinônimo de epigenética e isso se deve à grande difusão desse tema frente à sua constante e polêmica associação ao nome de Lamarck[7], [8], naturalista francês do século XVIII defensor da evolução biológica e que tem seu nome associado aos fenômenos do uso e desuso e da herança dos caracteres adquiridos[9]. Jablonka e Lamb, as principais teóricas da herança epigenética, denominaram a herança epigenética como a dimensão lamarckiana da evolução[10].
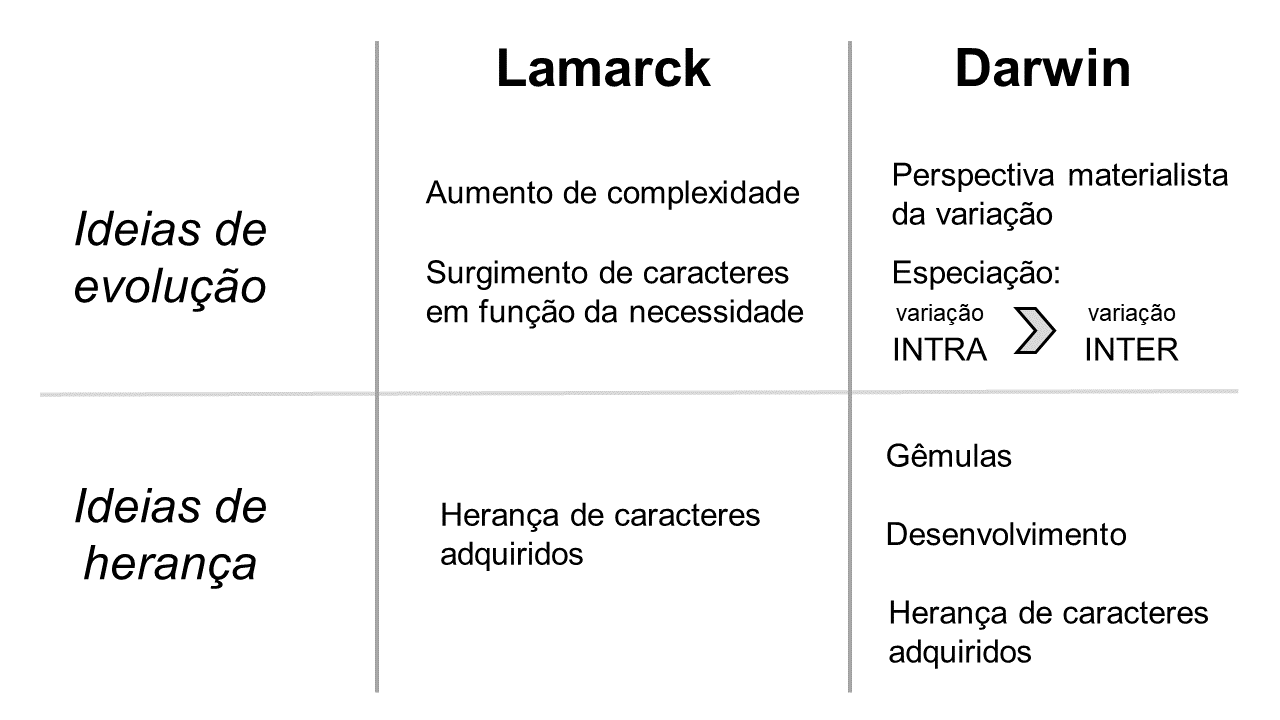
Lamarck (1744-1829) defendeu a ideia de que a evolução era uma tendência ao aumento do nível de complexidade dos organismos – exposta na sua primeira lei “tendência para o aumento da complexidade”. Do mesmo modo, ele tentou compreender quais mecanismos internos promoviam esse aumento de complexidade – sua segunda lei “surgimento de órgãos em função de necessidades que se fazem sentir e se mantém” – no sentido de responder às necessidades impostas pelo meio. Portanto, a teoria de Lamarck dizia que as variações novas – mudanças que ocorriam nos indivíduos ao longo de suas vidas – surgiam com função determinada, como uma resposta a uma necessidade do organismo. Desta forma, as variações, para Lamarck, eram necessariamente úteis à sobrevivência dos organismos e manutenção da espécie[11].
Segundo Darwin (1809-1882), a variação entre indivíduos dentro das populações era a realidade do mundo natural. Assim, a variação individual era pré-existente e, portanto, não atendia a necessidades impostas pelo meio. Dentro desta lógica, Darwin apresentou a especiação como um processo de transformação desta variação entre indivíduos (variação intrapopulacional) em variação entre grupos (variação interpopulacional), no qual os diferentes grupos divergiriam ao longo do tempo até se tornarem espécies biologicamente distintas. Deste modo, regredindo na linha do tempo, deduziu um ancestral comum para todas as espécies. Por outro lado, o avanço deste processo no tempo explicava a biodiversidade. Portanto, na teoria evolutiva darwiniana, a evolução acontecia sem propósito e não estava mais associada ao progresso, como na teoria de Lamarck, apenas à mudança. A metáfora da escada era substituída, assim, pela metáfora da árvore (FIGURA 2).
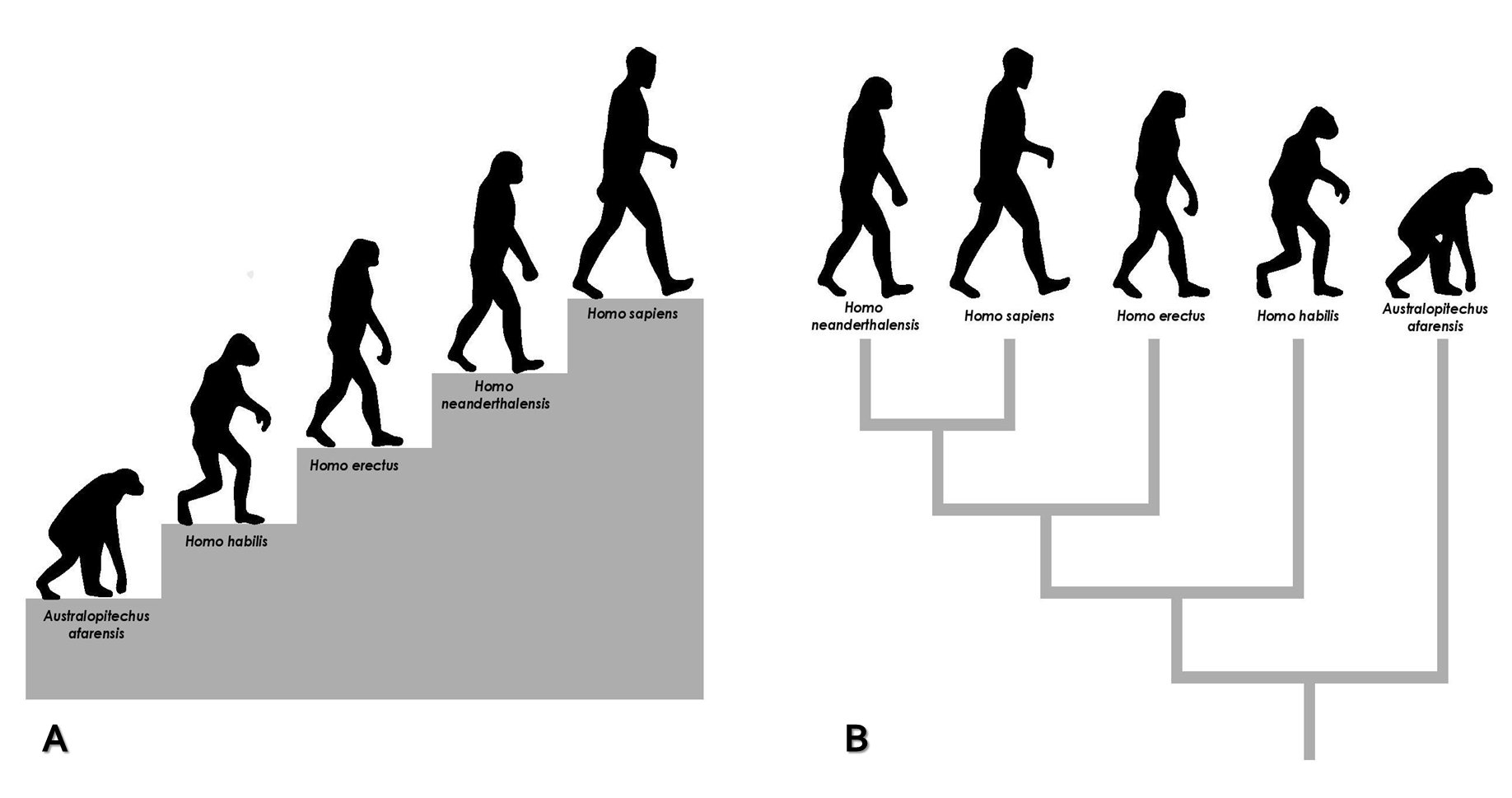
A epigenética faz parte do paradigma darwiniano, uma vez que aceita que a evolução é, basicamente, um processo de produção de variantes e escolha desses variantes (seja por seleção natural ou aleatóriamente por deriva genética). Dessa forma, ela não se associa a teoria de Lamarck e suas concepções finalistas de tendência ao aumento da complexidade e surgimento de variação como resposta a necessidades específicas. Sabe-se hoje que as variações presentes nas populações naturais têm origem em mutações genéticas ou epigenéticas e entende-se que, em última instância, essas mutações são cegas (ou seja, não tem nenhuma finalidade). Ainda que, ao longo do processo evolutivo, algo possa induzir a ocorrência de uma mutação (como, por exemplo, a exposição a uma fonte natural de radiação), esse evento não pode definir qual tipo de mutação irá ocorrer. A mudança não pode ser deduzida previamente, muito menos atender a qualquer “necessidade” do organismo. Portanto, o objetivo da comparação com Lamarck, na verdade, é fazer referência à lei da herança dos caracteres adquiridos, quarta (e mais popular) lei do naturalista francês. Segundo estas ideias, as mudanças que ocorressem no corpo de um indivíduo durante sua vida seriam passadas aos seus descendentes.
No entanto, a ideia de que características adquiridas durante o desenvolvimento somático poderiam ser transferidas para a geração seguinte não era exclusividade de Lamarck. A noção de herança do adquirido foi utilizada para explicar a semelhança entre descendentes e parentais desde a Grécia antiga, tendo sido Hipócrates de Cós II (460–370 a.C.), considerado o “pai da medicina”, um de seus primeiros proponentes. Mais que isso, essa noção fazia parte do senso comum da comunidade científica nos séculos XVIII e XIX. Lamarck, na realidade, não fez mais do que incluir uma ideia comum do seu tempo como pressuposto fundamental de sua teoria, sem discuti-la a fundo.
As leis “tendência para o aumento da complexidade” e “surgimento de órgãos em função de necessidades que se fazem sentir e se mantém” eram as verdadeiras novidades da teoria lamarckista e, entre elas e a epigenética, há pouca ou, talvez, nenhuma relação. Neste sentido, parece estranho que uma área de estudo do século XXI que tangencia a evolução e que, portanto, entende as variações como cegas seja formalmente denominada neo-lamarckista. Por outro lado, é possível encontrar muito mais semelhança entre as ideias da epigenética e outra teoria de herança baseada na herança de caracteres adquiridos, a teoria da pangênese de Darwin.
A “Hipótese Provisória da Pangênese”, publicada por Darwin no último capítulo de seu livro A Variação de Animais e Plantas sob Domesticação, recebeu esse nome por ser uma teoria não definitiva, na qual Darwin tentava responder as mais diversas questões a respeito da hereditariedade, assim como oferecer uma explicação para a origem e natureza da variação, indispensável para sua teoria evolutiva.
Resumidamente, a pangênese era baseada no pressuposto de que todo o corpo é responsável pela embriogênese. De acordo com esta teoria, cada parte do corpo seria capaz de produzir pequenas gêmulas que, por sua vez, carregariam a informação referente àquela região do organismo. Essas gêmulas, em dado momento, se acomodariam nos órgãos genitais. Durante a reprodução, os conjuntos de gêmulas dos parentais se reuniriam e, a partir desta reunião, seria gerado um novo indivíduo que expressaria características de ambos os pais (FIGURA 3). Um dos pressupostos da teoria era de que as gêmulas seriam continuamente produzidas pelo organismo ao longo de toda a vida do indivíduo, portanto, gêmulas de todas as épocas do desenvolvimento seriam armazenadas nos órgãos reprodutivos. Deste modo, as mudanças que ocorressem no corpo de um indivíduo ao longo de sua vida também passariam a ser codificadas em parte de suas gêmulas, podendo, portanto, ser passadas aos seus descendentes. Assim, Darwin forneceu um mecanismo consideravelmente sofisticado capaz de explicar a herança dos caracteres adquiridos.
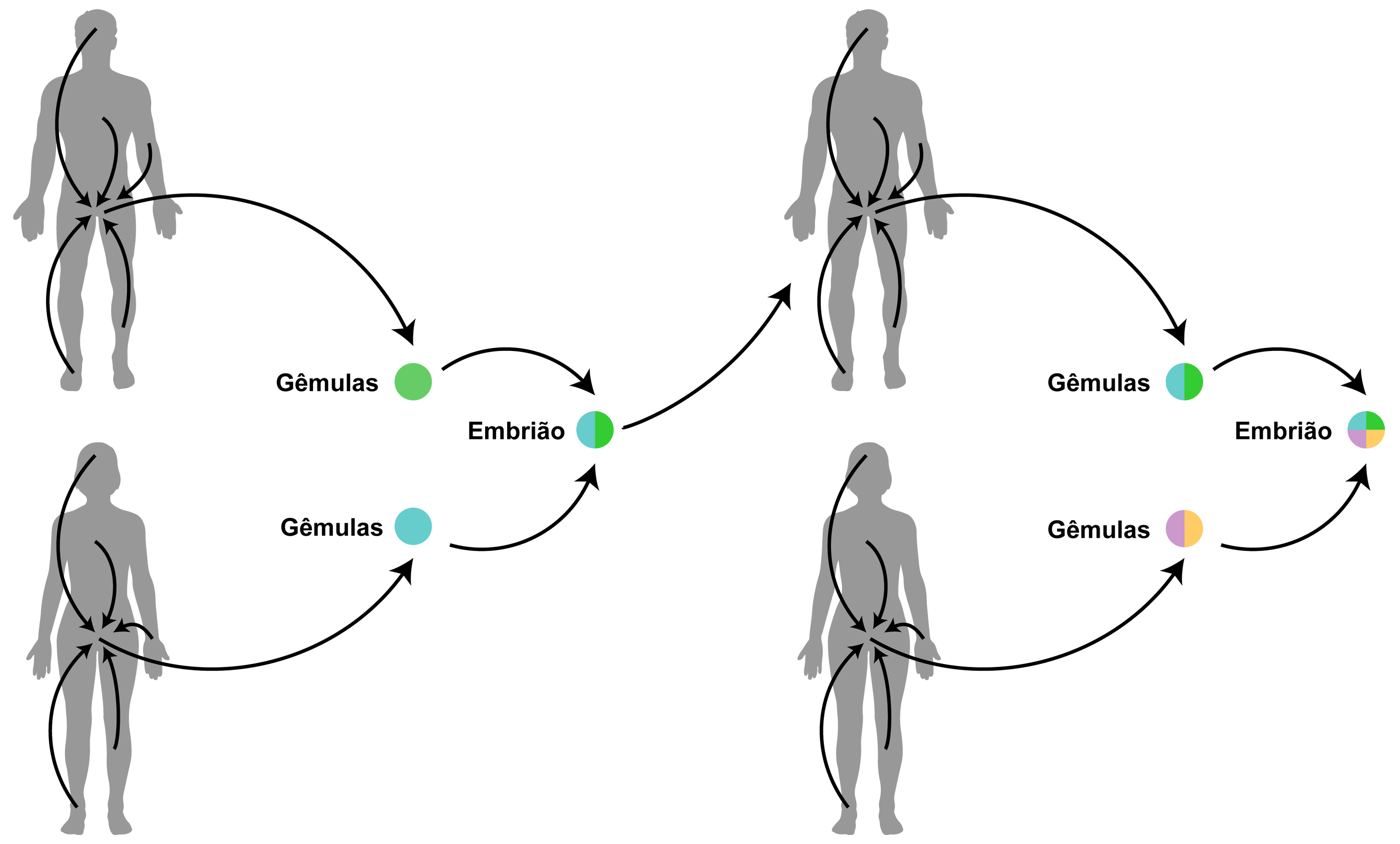
Embora ambos, Lamarck e Darwin, admitissem a ideia de herança do adquirido, a teoria de Lamarck era uma teoria evolutiva, que tinha como objetivo explicar como e porque os seres vivos se modificavam ao longo do tempo. Enquanto isso, a teoria da pangênese de Darwin era, antes de tudo, uma teoria da hereditariedade, preocupada com a origem da variação e como ela seria herdada. Mais que isso, era uma teoria com uma perspectiva do desenvolvimento (na época embriologia). Portanto, a “hipótese provisória da pangênese” de Darwin tem maior identidade com a herança epigenética e com os estudos da epigenética que a teoria de Lamarck (FIGURA 4).
As ideias sobre os mecanismos de herança desenvolvidas por Darwin na sua “Hipótese Provisória” eram boas respostas, na época, às perguntas que ele se propôs a responder e, mais que isso, compunham uma hipótese heurística, que poderia ser adotada como diretriz para o estudo da hereditariedade (Francis Galton, primo de Darwin, chegou a realizar experimentos com o intuito de testar a teoria da pangênese, da qual era simpatizante). Neste sentido, a pangênese não deveria ser relegada ao lixo da história. Sobretudo, a pangênese foi uma teoria que se propôs a explicar e reformar uma ideia, hoje central para a nova epigenética, aquela da herança dos caracteres adquiridos. Portanto, se alguma teoria deve ser historicamente relacionada à epigenética e ao fenômeno da herança epigenética, esta deveria ser a “Hipótese Provisória da Pangênese” de Darwin e não as ideias evolutivas de Lamarck.
* Os autores escrevem na variante brasileira do português.
[editar] Referências
- ↑ HOLLIDAY, R., Epigenetics: a historical overview. Epigenetics, 1:76–80, 2006.
- ↑ JABLONKA, E. & LAMB, M. J., The changing concept of epigenetics. Annals of the New York Academy of Science, 981:82-96, 2002.
- ↑ RAKYAN, V.K., et al., The marks, mechanisms and memory of epigenetic states in mammals. Biochemical Journal, 356:1-10, 2001.
- ↑ PRAY, L., Epigenetics: genome, meet your environment. Scientist, 18:14–20, 2004.
- ↑ WATTERLAND, R.A. & JIRTLE, R.L., <a class="a-link" target="_blank" href="https://www.ncbi.nlm.nih.gov/pmc/articles/PMC165709/">Transposable elements: targets for early nutritional effects on epigenetic gene regulation</a>. Molecular and Cellular Biology, 23:5293-5300, 2003.
- ↑ JABLONKA, E. & LAMB, M.J., Trangenerational epigenetic inheritance. In: Evolution, The Extended Synthesis. Pigliucii, M. & Müller, G.B. (eds.). MIT Press, Cambridge, pp. 137-174, 2010.
- ↑ HAIG, D., Weismann rules! OK? Epigenetics and the Lamarckian temptation. Biology and Philosophy 22:415-428, 2007.
- ↑ BURGGREN, W.W., Epigenetics as a source of variation in comparative animal physiology – or – Lamarck is lookin’ pretty good these days. Journal of Experimental Biology, 217:682-689, 2014.
- ↑ MAYR, E., Lamarck revisited. Journal of the History of Biology, 5:55-94, 1972.
- ↑ JABLONKA, E. & LAMB, M.J., Epigenetic Inheritance and Evolution: The lamarckian inheritance. Oxford University Press, Oxford & New York, 1995.
- ↑ RODRIGUES, R.F.C. & SILVA, E.P., Lamarck: fatos e boatos. Ciência Hoje, 48:68-70, 2011.
Recursos relacionados disponíveis na Casa das Ciências:
Criada em 12 de Fevereiro de 2018
Revista em 03 de Março de 2018
Aceite pelo editor em 4 de Outubro de 2018
