Breve guia de Paleontologia urbana
Referência : Silva, C. M., Pereira, S., (2023) Breve guia de Paleontologia urbana, Rev. Ciência Elem., V11(4):043
Autora: Carlos Marques da Silva e Sofia Pereira
Editor: João Nuno Tavares
DOI: [https://doi.org/10.24927/rce2023.043]

A geodiversidade urbana e os fósseis de rudistas.
O potencial educativo, de divulgação científica ou até, simplesmente, de fruição estética e lúdica da geodiversidade urbana é enorme. Os fósseis são elementos destacados da geodiversidade e, em contexto edificado, ocorrem integrados nos elementos de origem geológica que compõem a cidade, constituindo um recurso pedagógico riquíssimo, versátil e de fácil acesso[1]. São cada vez mais numerosos os aficionados da observação de fósseis urbanos e mais diversificadas e cativantes as atividades pedagógicas e de divulgação que neles se baseiam[2], [3], [4]. Já havia birdwatching, agora também temos fossilwatching, com a enorme vantagem de os fósseis serem bem menos esquivos!

Contudo, para que estes fósseis urbanos possam ser fruídos em pleno, é fundamental existir informação que permita identificá-los e enquadrá-los adequadamente. Com o presente trabalho inicia-se uma série de breves guias de paleontologia urbana[5] tendo como objetivo contribuir para esse aproveitamento, fornecendo informação concisa, clara e atualizada, que possa servir de base a atividades pedagógicas, de divulgação científica ou simplesmente de procura e observação descontraída dos inúmeros fósseis que ocorrem na cidade. A série inicia-se com, porventura, os fósseis mais comuns e mais óbvios em contextos urbanos nacionais, os somatofósseis[6] de bivalves rudistas, em virtude da sua associação geológica à rocha ornamental de aparato por excelência no nosso país[7], [8], [9] e não só[10], o calcário Liós (FIGURA 1).
Rudistas, o que são?
Os rudistas, nome pelo qual são comummente conhecidos os organismos da ordem Hippuritida[11], constituem um grupo biológico extinto que engloba alguns dos moluscos bivalves mais bem-sucedidos e mais extraordinários de sempre[12]. O grupo surgiu no Jurássico Superior, no Oxfordiano médio (há cerca de 158 milhões de anos, Ma)[13], [14], diversificou-se e floresceu durante o Cretácico, extinguindo-se no final do Mesozoico (há cerca de 66 Ma). Os rudistas foram, tudo indica, vitimados pelo mesmo evento de extinção em massa do final do Cretácico que varreu da face da Terra os dinossáurios não-avianos em ambientes continentais e as amonites em ambientes marinhos, entre muitos outros grupos biológicos extraordinários[15]STEUBER, T. et al., Catastrophic extinction of Caribbean rudist bivalves at the Cretaceous–Tertiary boundary, Geology, 30, 999–1002. 2002., [16], [17].
Foi Lamarck (1744–1829), proeminente naturalista francês, que individualizou e batizou o grupo em 1819[18]. O nome Rudista resulta do latim rudis, significando rude, em virtude do aspeto robusto e grosseiro da parede das conchas fossilizadas de muitos destes animais. Lamarck posicionou os rudistas no seio dos Conchifera, uma classe por ele definida que, para além dos organismos que hoje agrupamos nos bivalves, incluía também os braquiópodes. Ou seja, um grupo que reunia, em traços gerais, organismos possuidores de concha bivalve e desprovidos de cabeça. Seguindo esta lógica, na visão de Lamarck, os conchíferos constituíam uma entidade separada dos moluscos, do grupo que integrava os cefalópodes e os gastrópodes, por exemplo.
Contudo, para o sábio francês, a inclusão dos rudistas nos Conchifera não estava isenta de controvérsia. Para esta circunstância contribuiu o facto de, tal como definidos originalmente por Lamarck, “les Rudistes” serem um grupo heterogéneo. Isto é, para além de bivalves Hippuritida, tal como os entendemos hoje (e.g., géneros Sphaerulites e Radiolites), Lamarck incluiu nos seus rudistas géneros que atualmente são integrados nos braquiópodes (Discina, Crania) e até nos corais (Calceola)[19].
A dificuldade inicial em classificar adequadamente este grupo biológico, i.e., em posicioná- lo no seio da biodiversidade, resultava do facto de apenas ser conhecido do registo fóssil. Lamarck desconhecia que tipo de charneira e de ligamento uniria as valvas destes animais, ou até se possuiriam estas estruturas. Além disso, a morfologia das conchas (i.e., das partes duras, biomineralizadas, do corpo do animal) era muito diferente da dos restantes organismos bivalves conhecidos. Um aspeto, contudo, estava claro para Lamarck desde o início: os rudistas possuíam uma concha constituída por duas peças articuladas, por duas valvas, e isso— do seu ponto de vista— era suficiente para os incluir nos Conchifera.
Em tempos, os rudistas eram vistos como aberrantes, degenerescentes e votados ao fracasso, por serem extremamente distintos do que era familiar e por se terem extinguido no final do Mesozoico. Atualmente, pelo contrário, são considerados um grupo biológico inovador que experimentou uma diversificação notável, apresentando adaptações incomuns a ambientes particulares[20]. Além disso, extinguir-se faz parte do processo evolutivo. Não é— nem nunca foi— reflexo de insucesso biológico. Afinal, existir durante cerca de 90 milhões de anos (Ma), como o fizeram os rudistas, é obra. A esmagadora maioria dos grupos de organismos que desde há (pelo menos) 3,5 mil milhões de anos[21] existiram sobre a Terra já se extinguiu[22], [23] e os que atualmente subsistem, um dia, terão o mesmo destino.
Em que ambientes proliferavam?
Parafraseando livremente Louis Sullivan (1856–1924), o aclamado “pai dos arranha-céus” norte-americanos, em arquitetura, a forma é a materialização da função. Para Frank Lloyd Wright (1867–1959), autor do famoso edifício do Museu Guggenheim de Nova Iorque e ilustre discípulo de Sullivan, uma e outra estavam intimamente ligadas[24]. Pois bem, no mundo biológico, presente e passado, também. Ou seja, a morfologia dos organismos é indissociável da maneira como eles funcionam e de como se relacionam com o meio físico em que vivem, i.e., do seu modo de vida.
Por exemplo, os peixes têm barbatanas e nadam em ambientes aquáticos. As lagostas possuem apêndices locomotores articulados e deslocam-se autonomamente sobre o fundo marinho. Em suma, não há grupos biológicos desviantes, aberrantes ou degenerescentes. Há, isso sim, grupos que, por incluírem organismos que vivem (ou viveram) em condições fora do comum, apresentam formas e características que não nos são familiares, que são invulgares. Consequentemente, a existência de grupos biológicos apresentando características insólitas não denota afastamento de um padrão de perfeição biológico, mas sim diversidade de adaptações, maneiras diferentes de estar na vida. Essas características invulgares, nos rudistas ou em qualquer outro grupo biológico, não são defeito, são feitio. É esta linha de raciocínio que, juntamente com outras fontes de informação, nos permite entender como e em que ambientes os organismos do passado viviam, olhando para a sua morfologia preservada nos fósseis e interpretando-a em conformidade.
Assim, o que terá motivado a extraordinária morfologia destes bivalves mesozoicos? Em que ambientes ocorriam? De que modo se relacionavam com o meio em que viviam?
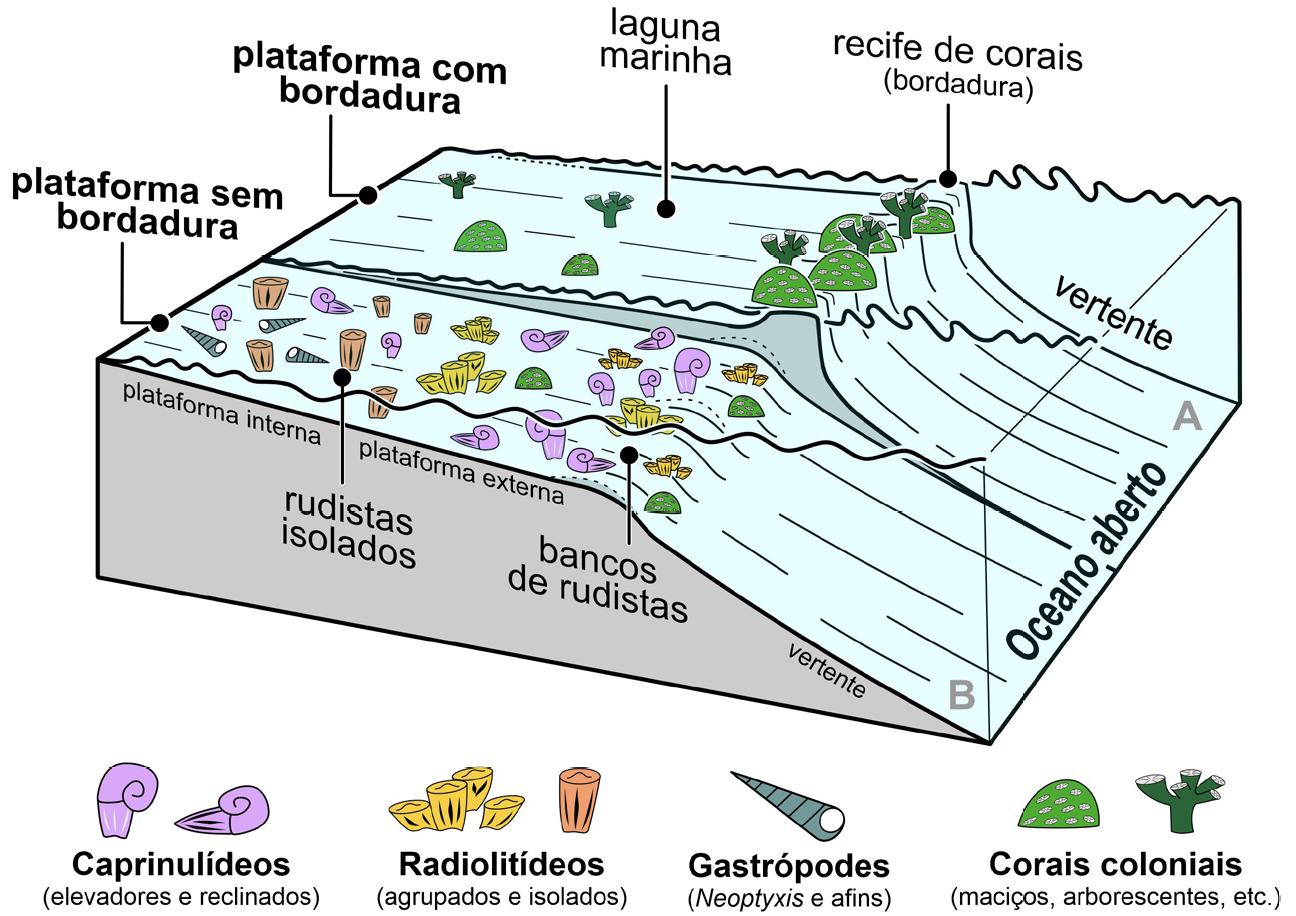
Os rudistas viviam em ambientes de plataforma carbonatada. Ou seja, habitavam em ambientes marinhos pouco profundos (preferencialmente, com cerca de 10 a 20 m de profundidade, não ultrapassando os 50 m[25]), bordejando a costa, em condições de águas quentes, tropicais a subtropicais, onde se depositavam predominantemente sedimentos carbonatados. A morfologia das plataformas carbonatadas é complexa, variando consideravelmente em função das condições em que se desenvolvem. A sua configuração é controlada quer por condicionantes geológicas, envolvendo a acumulação de maior ou menor quantidade de sedimento consoante as variações do nível do mar, quer ecológicas, relacionadas com a capacidade bioedificadora, geradora (ou não) de recifes, dos organismos que nelas habitam[26].
Ambientes de plataforma carbonatada, ainda que não exatamente iguais àqueles em que os rudistas habitavam, existem ainda hoje— por exemplo— em contexto tropical ao longo das costas da Flórida e nas Bahamas, no Oceano Atlântico, ou nas Maldivas, no Índico. Esses ambientes incluem, em termos muito gerais, uma laguna marinha de águas pouco profundas resguardada em maior ou menor grau da ondulação do mar aberto por uma bordadura bioedificada com expressão variável, podendo manifestar-se sob a forma de um recife (FIGURA 2A)). Do lado exterior da bordadura estende-se uma vertente exposta ao oceano. Nas plataformas carbonatadas cretácicas em que os rudistas predominavam, edificando bancos de rudistas, a barreira bioedificada não era tão destacada, tão proeminente, podendo até estar ausente, daqui resultando um ambiente marinho menos protegido da ondulação externa, i.e., mais exposto à influência do oceano (FIGURA 2B)). Entre um e outro tipo de plataforma carbonatada acima descrito existiam gradações, consoante os organismos bioedificadores que lá viviam (rudistas, corais, etc.) e o modo como se associavam, gerando barreiras recifais mais destacadas (FIGURA 2A)) ou menos expressivas (FIGURA 2B)).
Desde há cerca de 66 Ma[29], durante o Cenozoico e na atualidade, os principais organismos bioedificadores de recifes são os corais hexacoraliários (ou escleractiniários). No passado, no Paleozoico (entre os 539 e os 252 Ma), antes do aparecimento destes corais, esse papel foi desempenhado por organismos de vários outros grupos biológicos. Por exemplo, por arqueociatos, estromatoporóides e por corais tabulados e tetracoraliários, também conhecidos como corais rugosos, todos eles desaparecidos durante ou no final do Paleozoico. No Mesozoico, no Triásico e no Jurássico, os corais hexacoraliários, surgidos no Triásico, também tiveram um papel importante na construção de recifes. No Cretácico, contudo, os bioconstrutores por excelência foram os rudistas, o único grupo de moluscos a produzir autonomamente bioedificações significativas em contexto marinho em toda a história da vida na Terra.
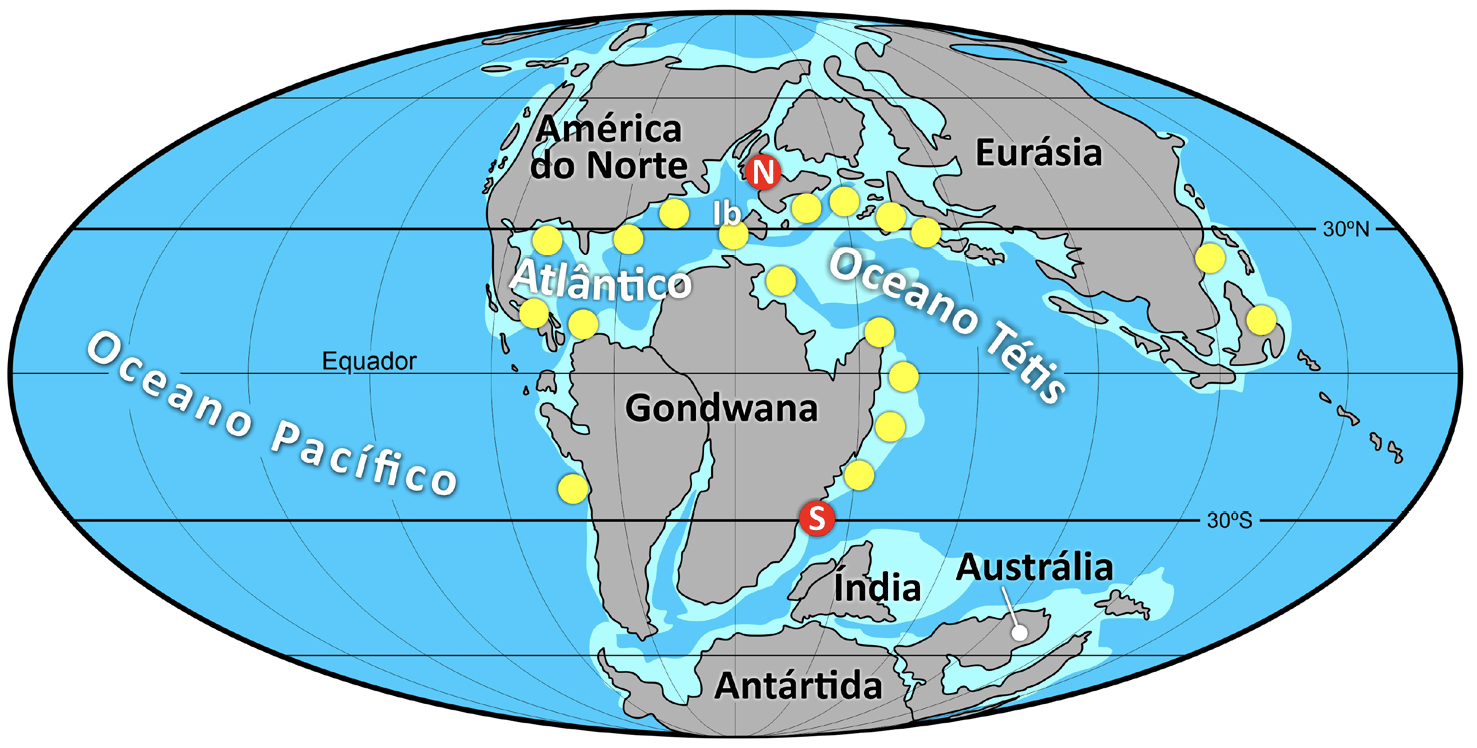
A partir de meados do Cretácico Inferior (Aptiano, 121 a 113 Ma) e, sobretudo, do Cenomaniano (Cretácico Superior, dos 100 aos 94 Ma) em diante, os rudistas proliferaram por toda a zona intertropical, dominando nos ecossistemas marinhos de plataforma carbonatada do antigo oceano Tétis, entretanto desparecido, e do jovem Atlântico (FIGURA 3).
Todavia, as estruturas produzidas pelos rudistas eram distintas das geradas pelos corais. O seu relevo acima do fundo marinho era diferente, menos destacado. Consequentemente, a configuração das plataformas carbonatadas em que proliferavam não era igual à daquelas onde o corais predominam (FIGURAS 2A) e B)).
Como viviam os rudistas?
Os rudistas eram organismos bentónicos, i.e., viviam no fundo marinho. Na sua maioria, ancoravam- se ao substrato por uma das valvas (i.e., eram organismos bentónicos sésseis ou fixos), convencionalmente denominada valva fixa, sendo a outra valva a livre. Mesmo aqueles rudistas que, na fase adulta, apenas assentavam sobre o sedimento (epibentónicos livres), iniciavam o seu crescimento fixados a algum tipo de substrato duro, por exemplo, a um fragmento de concha, i.e., a um bioclasto. Tal como a maioria dos bivalves, seriam organismos micrófagos suspensívoros: alimentavam-se de microrganismos microscópicos, sobretudo de fitoplâncton, e de diminutas partículas orgânicas existentes em suspensão na água[31].
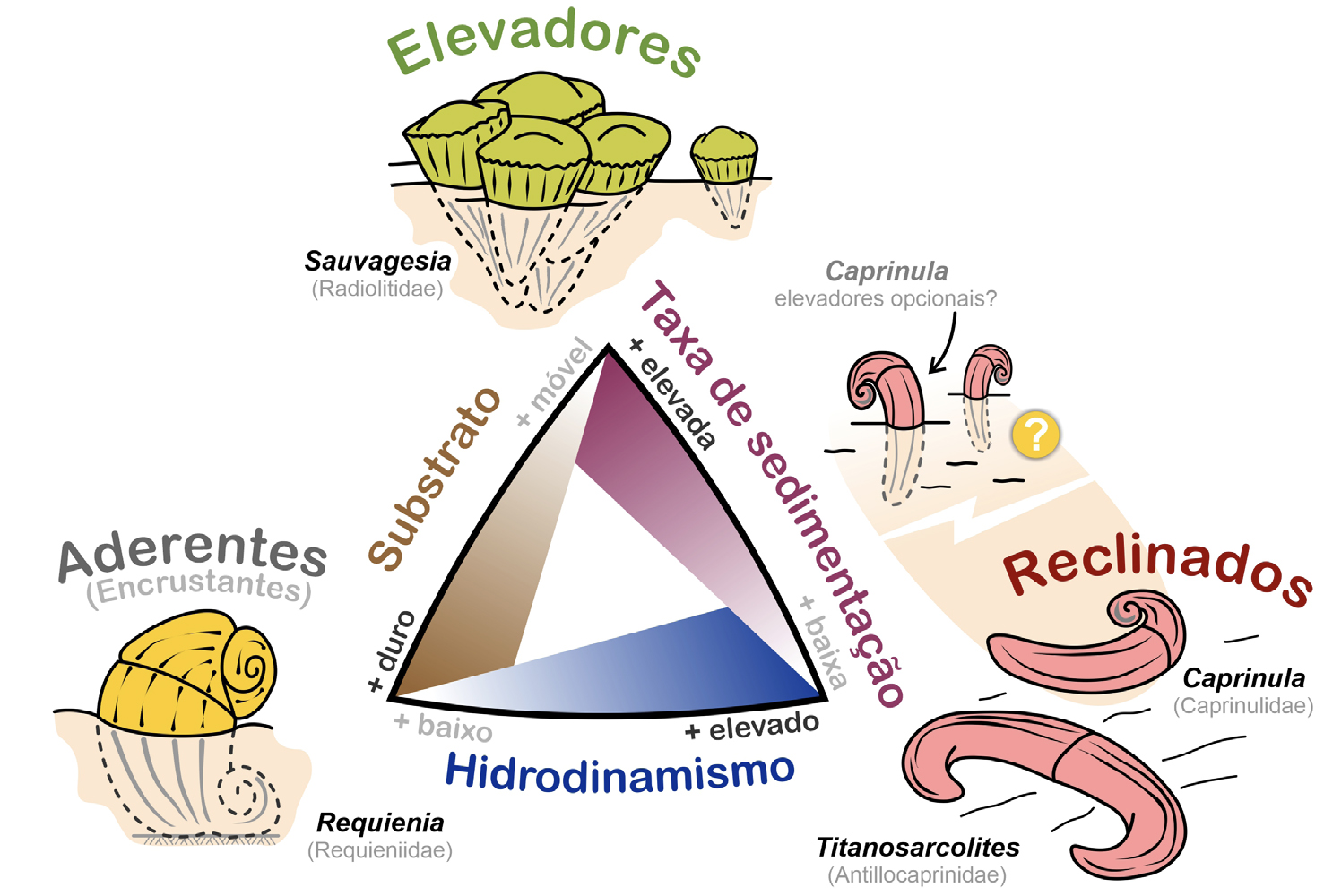
Tendo em conta o modo como os rudistas se relacionavam com o ambiente físico em que viviam— com o tipo de fundo (substrato), a velocidade de acumulação de sedimento (taxa de sedimentação) e a agitação das águas (hidrodinamismo)— distinguem-se três tipos morfológicos principais, ou seja, três morfótipos ecológicos: elevadores, reclinados e aderentes (ou encrustantes)[32], [33], [34], [35].
Nos rudistas elevadores, como o nome indica, a espessa valva fixa crescia para cima, elevando- se— ainda que pouco— acima da superfície do substrato (FIGURA 4). A sua valva livre era mais fina, aplanada ou arqueada, opercular, i.e., em forma de tampa. A estabilização do animal era conseguida pela ancoragem passiva da valva fixa no substrato móvel, como resultado da acumulação de sedimento à sua volta. Ou seja, estes animais viviam semienterrados no fundo. Frequentemente, os rudistas elevadores cimentavam-se uns aos outros, formando agregados de vários indivíduos, como se pode observar na FIGURA 1A). Os aderentes fixavam-se a substrato duro cimentando-se (FIGURA 4), aderindo ao fundo marinho pela valva fixa que se apresentava frequentemente algo espiralada. A estabilização da concha era conseguida pela ligação desta desta valva ao substrato. A valva livre podia ser mais aplanada, opercular ou espiralada. Os rudistas reclinados assentavam livremente no fundo (FIGURA 4), não se fixavam nem eram escorados pelo sedimento fino carbonatado. A imobilidade da concha resultava da circunstância de esta assentar no fundo horizontalmente, com ambas as valvas em contacto com o sedimento, de modo a gerar uma ampla superfície de estabilização. As valvas podiam ser arqueadas ou, em alguns casos, apresentar uma valva menos arqueada, subretilínea, e a outra espiralada. A forma arqueada e espiralada das valvas estabilizava a concha do bivalve no fundo marinho, impedindo que fosse facilmente virada pela agitação das águas.
Os rudistas elevadores, no contexto de plataforma carbonatada onde estes bivalves proliferavam, estavam mais frequentemente associados a ambientes de substrato móvel com taxa de sedimentação mais elevada e hidrodinamismo moderado a baixo. Os reclinados eram os mais bem-adaptados para ocupar substratos móveis em fundos com hidrodinamismo mais elevado e sedimentação baixa a nula. Por seu turno, os rudistas aderentes ocorriam mais frequentemente em ambientes com substrato mais estável a duro (bioclástico, i.e., com fragmentos de conchas, por exemplo) onde a sedimentação era baixa a nula e o hidrodinamismo baixo a moderado[36].
Estes morfótipos, contudo, não eram rígidos. Do mesmo modo que as condições ambientais eram variadas, também as adaptações dos organismos a elas eram diversificadas. Ocorriam termos intermédios entre os morfótipos básicos apresentados na FIGURA 4. Em alguns casos, os organismos de uma dada espécie podiam enquadrar-se em diferentes morfótipos ao longo do seu crescimento. Além disso, alguns rudistas seriam bastante adaptáveis às circunstâncias podendo, por exemplo, desenvolver-se facultativamente como reclinados ou como elevadores, consoante as condições ambientais[37]. Seria o caso de rudistas caprinulídeos como Caprinula, tão bem representados no Liós, ou Lioz (FIGURA 1B).
Recifes de rudistas ou bancos de rudistas?
Muitos organismos marinhos— tanto autotróficos como heterotróficos— são bioconstrutores ou bioedificadores. São organismos com esqueletos biomineralizados calcários, internos ou externos, que por via da sua capacidade de fixação ao substrato e/ou de agregação entre si geram estruturas diversificadas, com maior ou menor extensão e com relevo variável acima do substrato[39], i.e., acima do fundo marinho. Podem fazê-lo isoladamente ou em associação com organismos bioedificadores de outros grupos biológicos. Em determinadas circunstâncias, a estas bioedificações dá-se o nome de recifes.
Na linguagem quotidiana, um recife é um escolho ou um conjunto de rochedos submarinos localizados à flor da superfície do mar. Em contexto biológico e paleontológico, recife significa algo diferente. Assim, um recife é uma bioedificação combinando dois elementos fundamentais: primeiro, uma estrutura biogénica robusta, resistente que, segundo, se destaca claramente acima do fundo marinho, constituindo um relevo. Este tipo de construção está bem exemplificado, por exemplo, nos atuais recifes de corais tropicais.
Entre os rudistas, tal como nos bivalves que conhecemos da atualidade, havia organismos que viviam isolados, enquanto outros— de outras espécies— se agrupavam em grande número, por vezes aderidos entre si, formando extensas estruturas bioedificadas. Contudo, as bioconstruções de rudistas apresentavam-se, geralmente, mais dispersas, menos densas, que as dos corais recifais e, sobretudo, não se destacavam expressivamente acima do fundo, não originando relevos ou barreiras significativas.
O facto de os corais, para além de crescimento solitário, poderem apresentar crescimento colonial, i.e., de incluírem organismos clonais constituídos por vários indivíduos ligados fisicamente e funcionando integradamente como um todo, torna-os particularmente adequados à construção de recifes[40], [41]. Organismos de outros grupos com capacidade recifal, e.g. (por exemplo), os arqueociatos, estromatoporóides e os corais tabulados também apresentavam crescimento colonial. Além disso, na atualidade, os corais bioconstrutores (e por isso também denominados corais hermatípicos) possuem endossimbiontes fotossintéticos, i.e., possuem no seio dos seus tecidos moles organismos unicelulares capazes de realizar a fotossíntese. Estes organismos fotossintéticos pertencem ao grupo dos dinoflagelados e denominam-se coloquialmente zooxantelas. A sua associação aos corais recifais potencia a produção de esqueleto carbonatado dos corais, permitindo que as bioedificações por eles geradas se desenvolvam significativamente e se destaquem claramente acima do fundo.
Relativamente aos rudistas, contudo, estudos recentes[42] sugerem que — na esmagadora maioria dos casos — estes animais não possuiriam zooxantelas, i.e., que não seriam organismos fotossimbiontes. Segundo estes trabalhos, somente algumas espécies de rudistas (de radiolitídeos e de hipuritídeos) apresentavam conchas com aspetos morfológicos consentâneos com a exposição do bordo do manto, parte do corpo mole do bivalve, à luz solar, podendo ser interpretados como possíveis evidências— ainda que indiretas— da presença de organismos endobiontes fotossintéticos. Além disso, os rudistas, sendo bivalves, eram organismos solitários. Num organismo solitário— e.g., nos seres humanos – cada ser vivo corresponde a apenas um indivíduo capaz de funcionar autonomamente. Este tipo de crescimento solitário, aclonal, dos rudistas torna-os mais vocacionados para a ocupação oportunista do ambiente por um grande número de indivíduos da mesma espécie ou de poucas espécies. Estes bivalves tiravam partido das circunstâncias ecológicas disponíveis, mais propícias para eles que para os restantes grupos biológicos, de modo mais rápido e eficaz. Ou seja, aproveitavam as oportunidades (ecológicas) existentes, daí serem oportunistas. É o que se passa na atualidade com as ostras, um outro tipo de bivalves gregários. Elas ocupam fundos estuarinos com grande número de indivíduos, cobrindo áreas extensas, por vezes cimentando-se umas às outras, mas não se destacando significativamente acima do substrato. As ostras geram bancos de ostras, não edificam recifes.
Estruturas como as geradas pelos corais hermatípicos, elevando-se claramente acima do fundo, são denominadas suprastratais[43], [44] (FIGURA 5A)). As bioedificações mais frequentemente geradas pelos rudistas, semienterradas no sedimento que constitui o fundo, não se destacando significativamente acima dele, acompanhando a formação dos estratos, são classificadas como constratais (FIGURA 5B)). Ou seja, os rudistas não geravam recifes suprastratais, similares aos dos corais, mas sim bancos constratais[45], [46], [47], mais aproximados do relevo das atuais bioedificações de ostras.
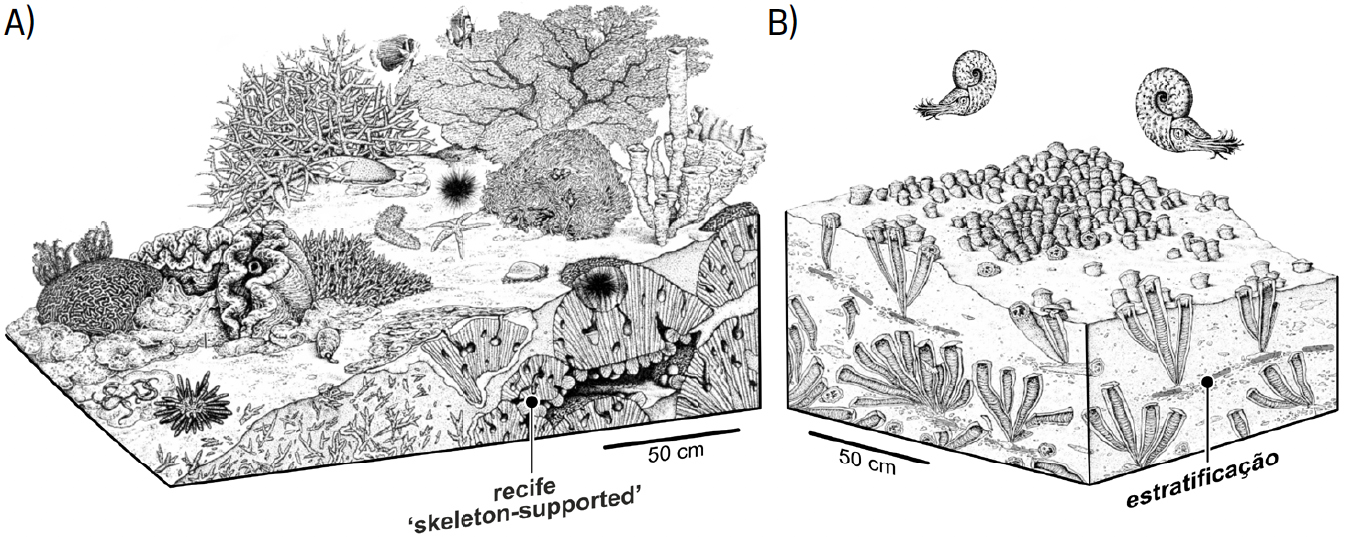
A classificação das bioconstruções pode variar de autor para autor. Como frequentemente acontece em ciência (e não só) diferentes investigadores têm perspetivas distintas sobre o mesmo tema, perspetivas essas incorporando vantagens, mas também padecendo de deficiências. Não há modelos perfeitos! Assim, alguns autores[50] enquadram as bioconstruções de rudistas no âmbito dos recifes, pois consideram todas as bioconstruções como sendo recifes de algum tipo. Contudo, mesmo nestas classificações, as estruturas geradas por rudistas são vistas como distintas das dos corais hermatípicos. As dos rudistas são incluídas na categoria de recifes sustentados pela matriz (i.e., matrix-supported reefs na terminologia anglo-saxónica), enquanto os recifes de corais são atribuídos à categoria dos recifes sustentados pelos elementos esqueléticos dos seus construtores (skeleton-supported reefs).
Nos recifes sustentados pelo sedimento os organismos bioconstrutores não se encontram em contacto uns com os outros extensivamente, gerando por isso aglomerados de baixa densidade. Daqui resulta que nos fundos marinhos ocupados por estas estruturas haja mais sedimento que organismos bioconstrutores, permitindo inclusive que se forme estratificação no seu seio, i.e., são constratais (FIGURA 5B)). Como resultado, a extensão destes recifes aglomerados (cluster reefs na terminologia anglo-saxónica) pode ser significativa, mas o seu relevo é reduzido. Pelo contrário, nos recifes construídos por corais toda a armação recifal resulta do crescimento em contacto dos diferentes organismos (frame reefs)[51]. Nestes casos, o suporte esquelético do recife é muito mais denso e resistente, destacando-se claramente acima do fundo e formando uma barreira, i.e., são estruturas suprastratais (FIGURA 5A)).
A capacidade bioedificadora dos rudistas variou ao longo da sua história. Durante o Jurássico Superior, do Oxfordiano médio ao Titoniano (dos 158 aos 145 Ma), não formavam aglomerados, surgindo dispersos em associação com outros organismos. Ocorriam frequentemente com corais, em recifes coralígenos onde, juntamente com outros bivalves, gastrópodes, equinodermes, etc., eram um dos vários elementos das comunidades marinhas de então. Mais tarde, a partir de meados do Cretácico Inferior (Aptiano), há cerca de 121 a 113 Ma, e sobretudo no Cretácico Superior, a partir dos 100 Ma (Cenomaniano), os rudistas dominaram os ecossistemas marinhos de plataforma carbonatada do antigo oceano Tétis e do jovem Atlântico (FIGURA 3). Nesses ambientes do final do Mesozoico os rudistas tornaram-se os principais organismos bioconstrutores.
Muito embora os rudistas gerassem, mais frequentemente, bancos constratais mono ou paucispecíficos, i.e., constituídos por uma ou por poucas espécies (FIGURA 6), a verdade é que também podiam coexistir com corais, contribuindo para a construção de recifes coralígenos mistos de corais e rudistas complexos e diversificados. A dinâmica dessa coexistência era determinada pelas condições ambientais. Há exemplos dessas bioedificações mistas no registo geológico das Caraíbas[52] e no do extremo ocidental do antigo oceano Tétis, em Espanha[53], [54] e em Portugal[55], por exemplo.

Em plataformas carbonatadas do Cretácico Inferior do Arizona (EUA) com cerca de 113 a 100 Ma (Albiano), por exemplo, corais e rudistas coabitavam frequentemente[56], [57], [58]. Contudo, os corais proliferavam quando a profundidade era maior e o influxo sedimentar externo menor, enquanto os rudistas prevaleciam em águas menos profundas, nos períodos em que o acarreio de sedimento era mais intenso. Porém, no registo geológico do Cenomaniano (Cretácico Superior, 100 a 94 Ma) da região de Lisboa, pelo menos em contexto de geodiversidade urbana, no calcário Liós, não é comum encontrar fósseis de corais, nomeadamente de corais coloniais, associados a aglomerações de rudistas[59]. No Liós, mais frequentemente, encontram-se associações mono ou paucispecíficas de rudistas radiolitídeos (FIGURA 1A)) e caprinulídeos (FIGURAS 1B) e 6).
Os rudistas caprinulídeos predominavam em zonas mais externas das plataformas carbonatadas, em condições de agitação marinha mais elevada, mais expostas à influência do mar aberto[60], [61]. Mas também podiam ocorrer em áreas mais interiores, em condições mais abrigadas, e inclusive na vertente exterior da plataforma, onde por vezes ocorriam em associação com corais, como registado para o Cretácico Superior de França[62]. Os caprinulídeos não formavam bioconstruções com estrutura rígida (frame reefs), pois não se cimentavam uns aos outros. Uma vez que eram predominantemente reclinados (ainda que também haja evidências de caprinídeos e de caprinulídeos elevadores[63], [64], [65], [66]) geravam bancos extensos, mas algo dispersos, pouco densos[67], [68], [69].
Pelo contrário, a maioria das bioconstruções de rudistas elevadores— e.g., de radiolitídeos— desenvolver-se-ia em ambientes de plataforma pouco profundos e protegidos, em contexto de plataforma interna mais abrigada, onde o hidrodinamismo (i.e., a agitação das águas) seria menos intenso[70], [71]. Em zonas ainda mais interiores da plataforma carbonatada, os radiolitídeos ocorriam isoladamente, coabitando com gastrópodes do grupo das nerineas, e.g., Neoptyxis, e dos acteonelídeos, tais como Acteonella (FIGURA 7A)), bem como com crustáceos e esponjas, estes dois últimos grupos representados nas associações fossilíferas por icnofósseis[72] (FIGURAS 7B) e 7C)).

Estas associações, incluindo ainda rudistas caprinulídeos, estão patentes em contexto urbano, e.g., nos calcários ornamentais Amarelo de Negrais (FIGURA 8) e Encarnado da Pedra Furada de idade cretácica superior (Cenomaniano) de Almargem do Bispo e de Montelavar (Sintra), respetivamente.
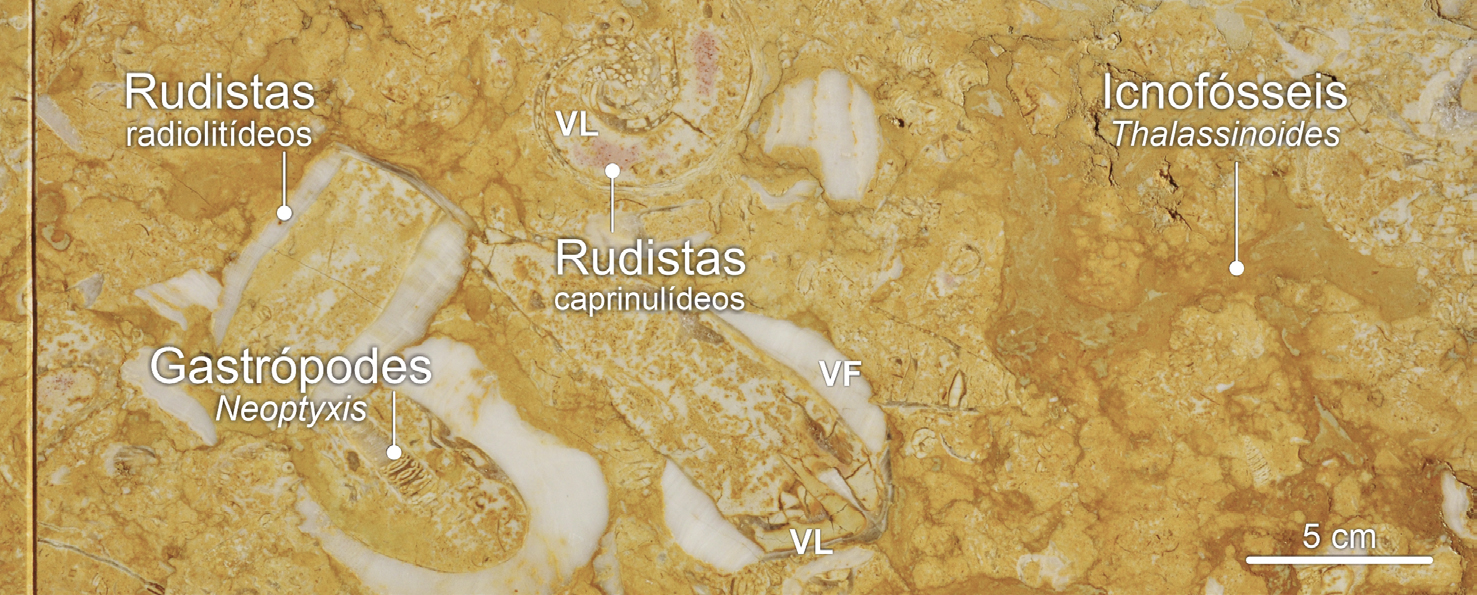
O Liós, os fósseis de rudistas e a cidade.
Nos Estados Unidos da América, do mesmo modo que, para efeitos de divulgação científica, de preservação da biodiversidade ou de afirmação de identidade cultural, muitos estados designaram uma ave estadual (state bird no original em inglês), ou uma flor estadual (state flower), também indicaram— tendo a geodiversidade em mente— um state fossil[75], [76]. Em Portugal esta abordagem (ainda) não existe. Mas se existisse, o fóssil representativo da região de Lisboa seria seguramente um rudista! Ocorrendo em rochas de idade cretácica aflorando descontinuamente desde a região de Lisboa até à Figueira da Foz, os fósseis de rudistas encontram-se também um pouco por todo o lado em muitas cidades nacionais associados— sobretudo— ao calcário Liós. Uma vez vistos, não mais deixamos de os reconhecer em fachadas, pavimentos, monumentos e até nas escadas e nas bancadas de cozinha de nossas casas, sobretudo nas mais antigas (FIGURA 1).
Na mesma linha de pensamento do fóssil regional, a rocha local mais emblemática de Lisboa é, sem sombra de dúvida, o Liós. Este nome é a designação dada na indústria das rochas ornamentais ao calcário cretácico (Cenomaniano, cerca de 100 a 94 Ma) de coloração variada, mas tipicamente bege, microcristalino, afetado por estilólitos[77], bioclástico, ostentando fósseis evidentes de rudistas, que ocorre na região de Lisboa— Mafra[78]. O Liós, pelas suas características estéticas e físico-mecânicas, nomeadamente pela sua durabilidade, é uma rocha ornamental muito apreciada. São distinguidas diversas variedades, consoante o seu aspeto geral, coloração e conteúdo fóssil. Liós, Liós Creme, Abancado (também, por vezes, referido como Encarnado de Morelena[79] e Encarnadão), St. Florient Rose e Azulino, são atualmente algumas das mais populares. Estas variedades podem ser pesquisadas online no Catálogo de Rochas Ornamentais Portuguesas, ORNABASE[80], do Laboratório Nacional de Energia e Geologia (LNEG).
Não obstante aflorar apenas na região de Lisboa-Sintra, o calcário Liós, por ser uma rocha muito valorizada, de prestígio, explorada já desde tempos romanos[81], pode ser encontrada em edificações por todo o país. Aliás, há Liós disseminado por todo o mundo de influência cultural portuguesa[82], [83], [84]. Recentemente, foi noticiado que o padrão português implantado por Diogo Cão em 1485 na Costa dos Esqueletos foi devolvido à Namíbia pelo Deutsches Historisches Museum, para onde havia sido levado em 1893, na sequência da ocupação daquele território sul-africano pela Alemanha[85]. Esse padrão, atualmente à guarda das autoridades namibianas, foi seguramente talhado em Liós da região de Lisboa. Curiosamente, o Liós— presente em muitos pavimentos— não é usado na característica calçada portuguesa[86].

O Liós é um recurso geológico tão importante, tão emblemático, que transpôs a fronteira da História Natural e da Arquitetura para o domínio, por exemplo, das artes decorativas[87], [88]. Podemos encontrar representações de Liós com rudistas em inúmeras pinturas de fingimento, trompe l’oeil, tanto sobre estuque, como sobre madeira e azulejos, em igrejas, conventos e palácios lácios e até em casas de habitação de finais do séc. XIX e inícios do séc. XX (FIGURA 9A) e B)). A relevância económica e, acima de tudo, cultural deste recurso geológico nacional valeu-lhe, em 2019, a classificação como Global Heritage Stone Resource pela UNESCO e a IUGS. E com o Liós, para a cidade, vêm os fósseis de rudistas.
Apesar do destacado valor económico e cultural do Liós e de outras rochas ornamentais, a sua presença na cidade não está a salvo de ameaças. O desordenamento do espaço construído e a falta de manutenção das estruturas edificadas são propícios a ações que degradam, mutilam e tornam ilegível muita da geodiversidade presente nos espaços edificados (FIGURA 9C)). Manifestações tipicamente urbanas como grafitos e tags, bem como a publicidade excessiva e desregrada, contam-se entre as mais gravosas para a adequada conservação do edificado e para um usufruto pleno da geodiversidade urbana.
Que rudistas se encontram na cidade e como os identificar?
Em Portugal, em contexto de geodiversidade urbana (que, nunca é demais lembrar, não deve ser confundido com ocorrências naturais, nem a elas igualado), a maioria dos fósseis de rudistas surge nas diversas variedades do calcário Liós. Também se encontram fósseis de rudistas associados, por exemplo, ao calcário Amarelo de Negrais e ao Encarnado da Pedra Furada, também de idade cenomaniana, igualmente da região de Sintra (FIGURAS 8 e 14). Nestas rochas ornamentais estão bem representados dois grupos de rudistas: os radiolitídeos (família Radiolitidae), sobretudo representados pelo género Sauvagesia, mas também por Radiolites s.l., e os caprinulídeos (família Caprinulidae), representados por Caprinula. É importante ter-se em consideração que a identificação de fósseis em corte em contexto urbano, em fachadas e pavimentos, não é uma ciência exata. Esta é uma abordagem expedita tendo em vista a divulgação científica e o usufruto dos fósseis que ocorrem na cidade, não uma análise paleontológica convencional em condições laboratoriais ótimas. É a abordagem— e a identificação— possível, não a ideal.
Radiolitídeos e caprinulídeos apresentam conchas com formas bastante distintas (FIGURA 10), correspondendo a morfótipos diferentes (FIGURA 4). Os radiolitídeos apresentam valva fixa cónica (a que estava ancorada no sedimento e que alojava no seu interior o corpo mole do bivalve) e valva livre opercular aplanada ou levemente arqueada (FIGURA 10A)).
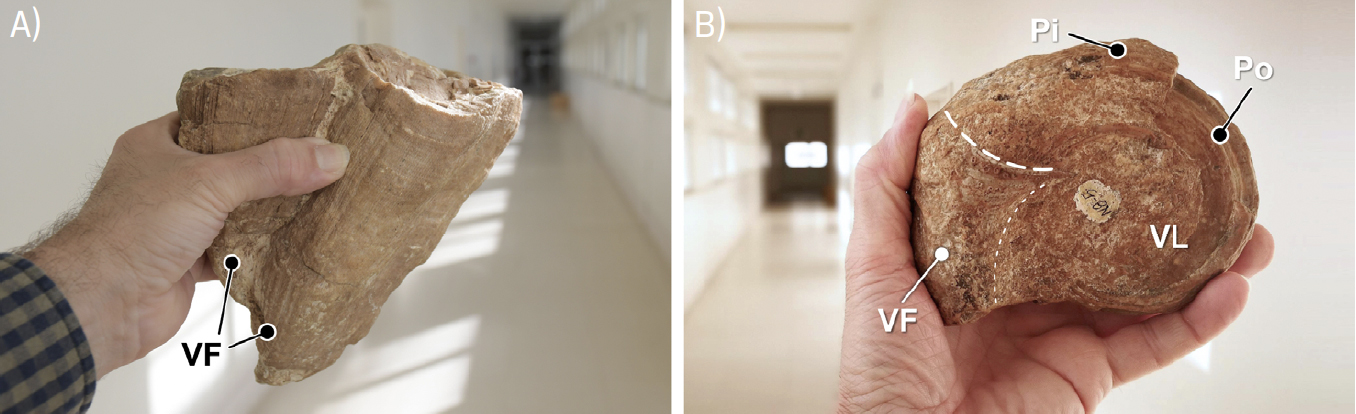
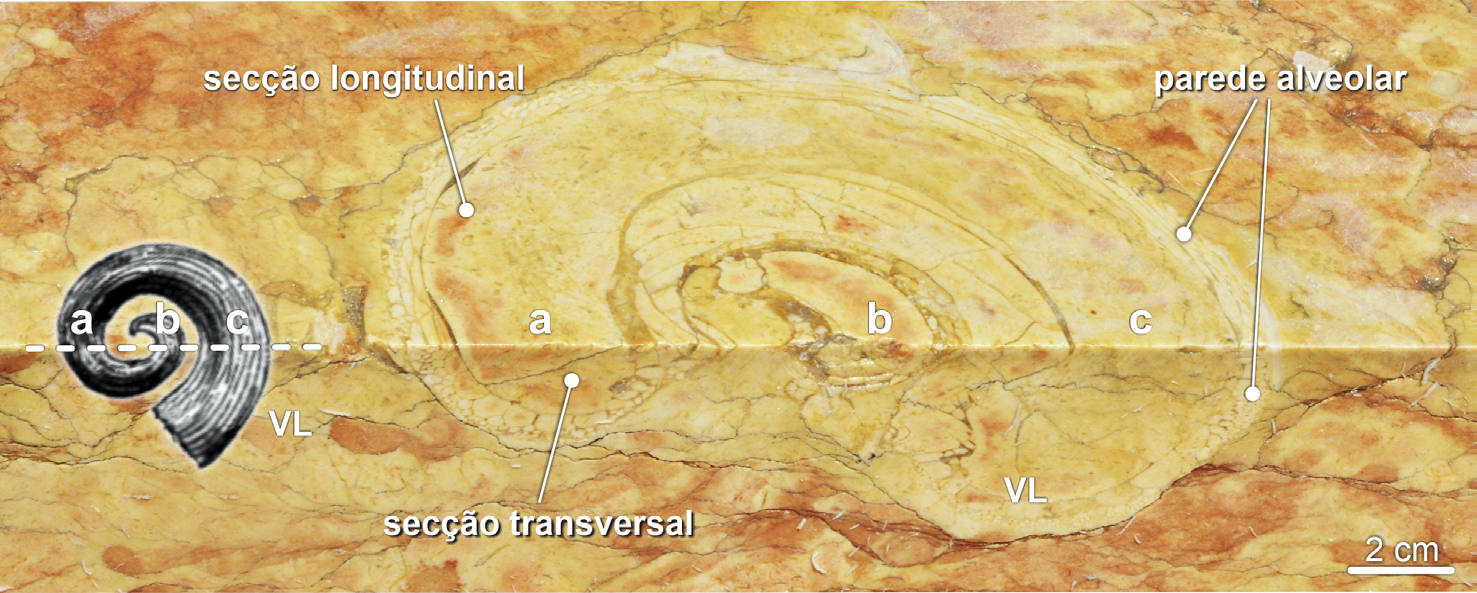
Por vezes, como sucedia com vários outros rudistas, a valva fixa, cónica, dos radiolitídeos ostentava divisórias interiores horizontais, as tábulas. Estas estruturas são mais comuns nos rudistas do grupo dos Radiolites, patentes no calcário Amarelo de Negrais. Note-se que, em Radiolites s.l., o perfil da valva fixa (mais evidente em cortes longitudinais) é bastante mais sinuoso que em Sauvagesia (FIGURA 7C) e 11). As tábulas separavam as partes desocupadas do interior da concha, mais antigas, da porção ocupada pelo corpo mole do animal (FIGURA 11B)).
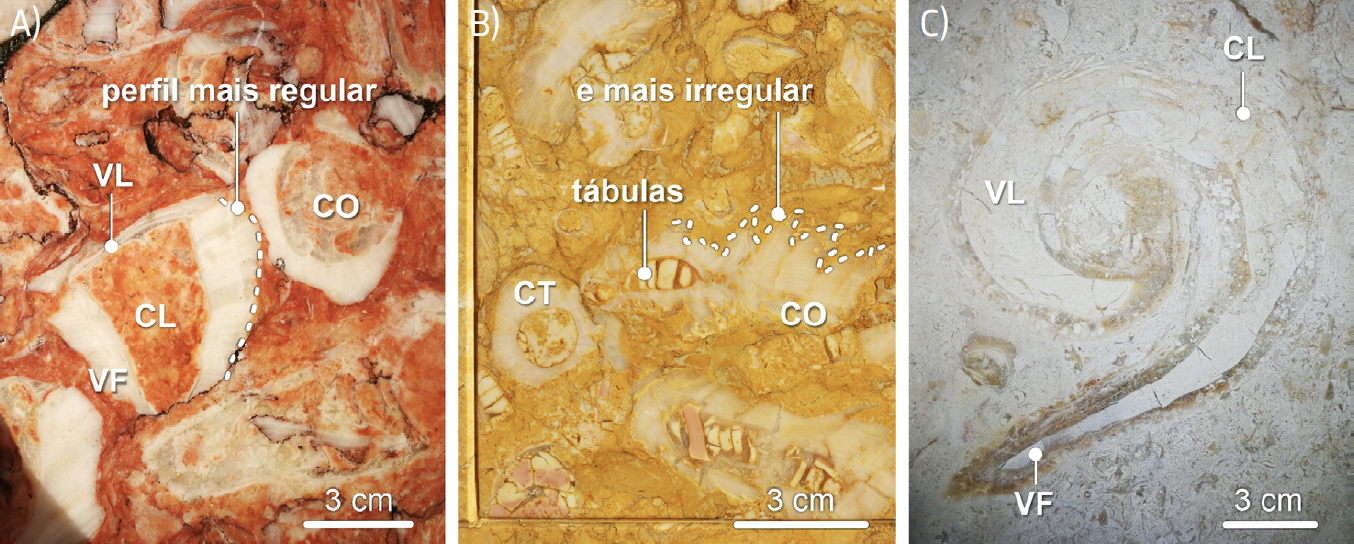
Os radiolitídeos eram rudistas elevadores, gregários, podendo surgir cimentados uns aos outros (FIGURAS 1A) e 10A)). De notar que as conchas dos rudistas, tratando-se de bivalves, tendem a desarticular-se após a morte do organismo. Assim, mais frequentemente se encontram fossilizadas valvas isoladas das conchas destes animais, sobretudo valvas fixas, mais espessas e resistentes. Contudo, ainda que menos comuns, também ocorrem fósseis de conchas com as valvas articuladas (e.g., FIGURAS 13B) e 13C)). Os caprinulídeos ostentam valva “fixa” cónica, por vezes arqueada, de dimensão variável, e valva livre espiralada. Consoante as condições ambientais, podiam comportar-se como rudistas reclinados— mais frequentemente— ou elevadores[89] (FIGURA 4). Os caprinulídeos, ainda que gregários, viviam isolados, nunca se cimentando uns aos outros, formando associações de baixa densidade. Todos estes rudistas proliferavam em ambientes de plataforma carbonatada (FIGURA 2). Contudo, os radiolitídeos predominariam em zonas mais internas, de hidrodinamismo mais baixo, enquanto os caprinulídeos proliferariam em zonas mais exteriores, mais expostas, com agitação das águas mais elevada, associadas à margem externa da plataforma[90], [91] (FIGURA 2).
Em contexto urbano, em fachadas e pavimentos, expostos em corte, a distinção mais óbvia entre rudistas radiolitídeos e caprinulídeos reside na estrutura da parede das conchas. Os radiolitídeos ostentam concha espessa, maciça, com estrutura celuloprismática fina (FIGURA 12A)), nem sempre evidente. Os caprinulídeos apresentam concha mais delgada, com estrutura alveolar, bem patente no Liós. Em corte transversal, a parede da concha dos caprinulídeos apresenta, na periferia, canais paliais piriformes, estreitos, alongados, com contorno em pera— daí a designação piriformes— ou em gota. A zona mais interior da parede exibe canais poligonais, mais grosseiros, com contorno mais anguloso (FIGURA 12B)).
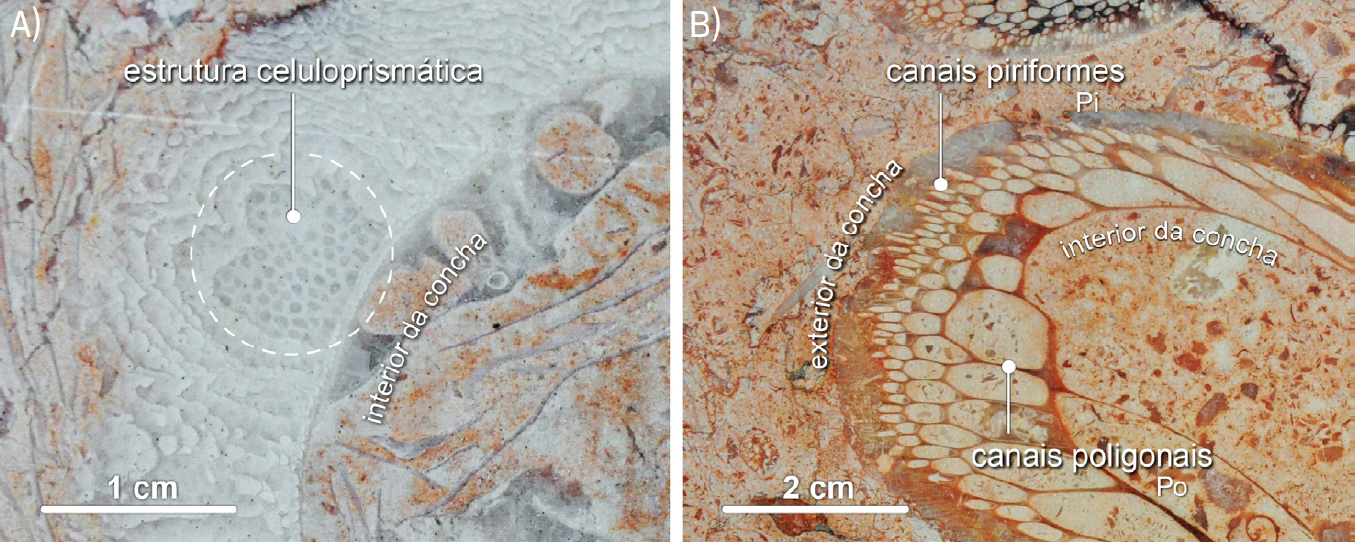
O género Caprinula, bem representado em formações cretácicas nacionais[92], [93], era tradicionalmente incluído na família dos rudistas caprinídeos (Caprinidae) em virtude de, por exemplo, tanto Caprina como Caprinula apresentarem estrutura da concha ostentando canais paliais[94]. Contudo, estudos recentes[95], [96] revelaram a natureza heterogénea da família Caprinidae tal como era anteriormente entendida. Daí que vários géneros de rudistas, incluindo Caprinula, tenham sido individualizados na família Caprinulidae[97], [98], os caprinulídeos.
Ocorrendo em contexto urbano em corte, na superfície de paredes e em pavimentos, para identificar e interpretar corretamente os fósseis de rudistas há que ter em conta a orientação dos cortes (transversal, longitudinal, etc.), quer para os radiolitídeos (FIGURA 13) quer para os caprinulídeos (FIGURA 14).
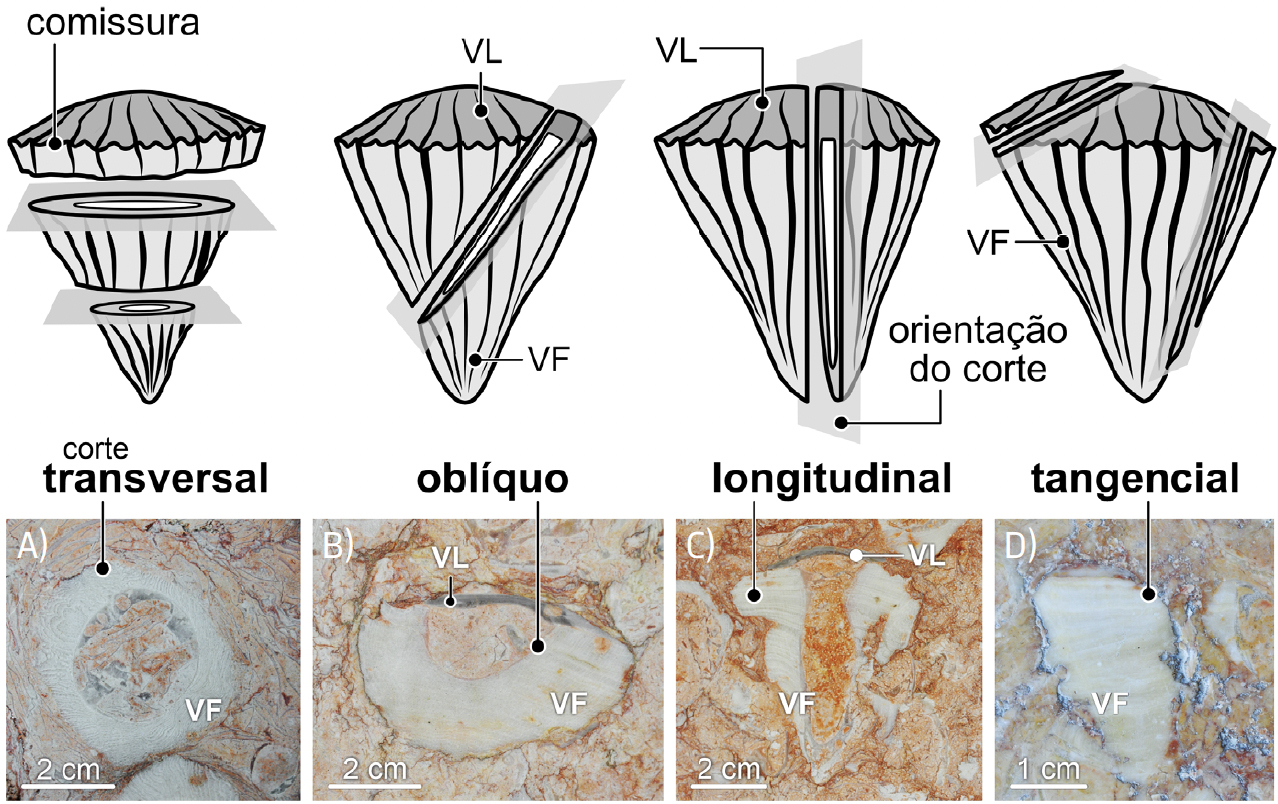
Curiosamente, a investigação científica dos rudistas também se faz, em grande parte, estudando os seus fósseis em corte. Isto deve-se ao facto de, por um lado, os rudistas ocorrerem mais frequentemente em rochas carbonatadas, em calcários, sendo difícil extraí- -los da pedra e, por outro lado, por os aspetos da sua morfologia mais diagnósticos, mais informativos, se encontrarem na parede ou no interior das conchas, não à sua superfície. Além disso, tal como com as ostras na atualidade, a forma exterior das conchas dos rudistas (inclusive no seio de uma mesma espécie) podia variar significativamente de acordo com as condições ambientais. Ou seja, tal como as ostras, os rudistas eram organismos polimórficos, i.e., podiam apresentar formas algo diferentes, dentro de limites conhecidos, mas amplos.
No calcário Liós, os fósseis de rudistas mais evidentes são os cortes transversais das valvas fixas de radiolitídeos. São frequentemente descritos pelo público em ações de observação de fósseis na cidade como “rodelas de ananás” ou “ovos estrelados” devido ao facto de serem arredondados, apresentando parede espessa e ampla cavidade interna (FIGURAS 1A) e 13A)).
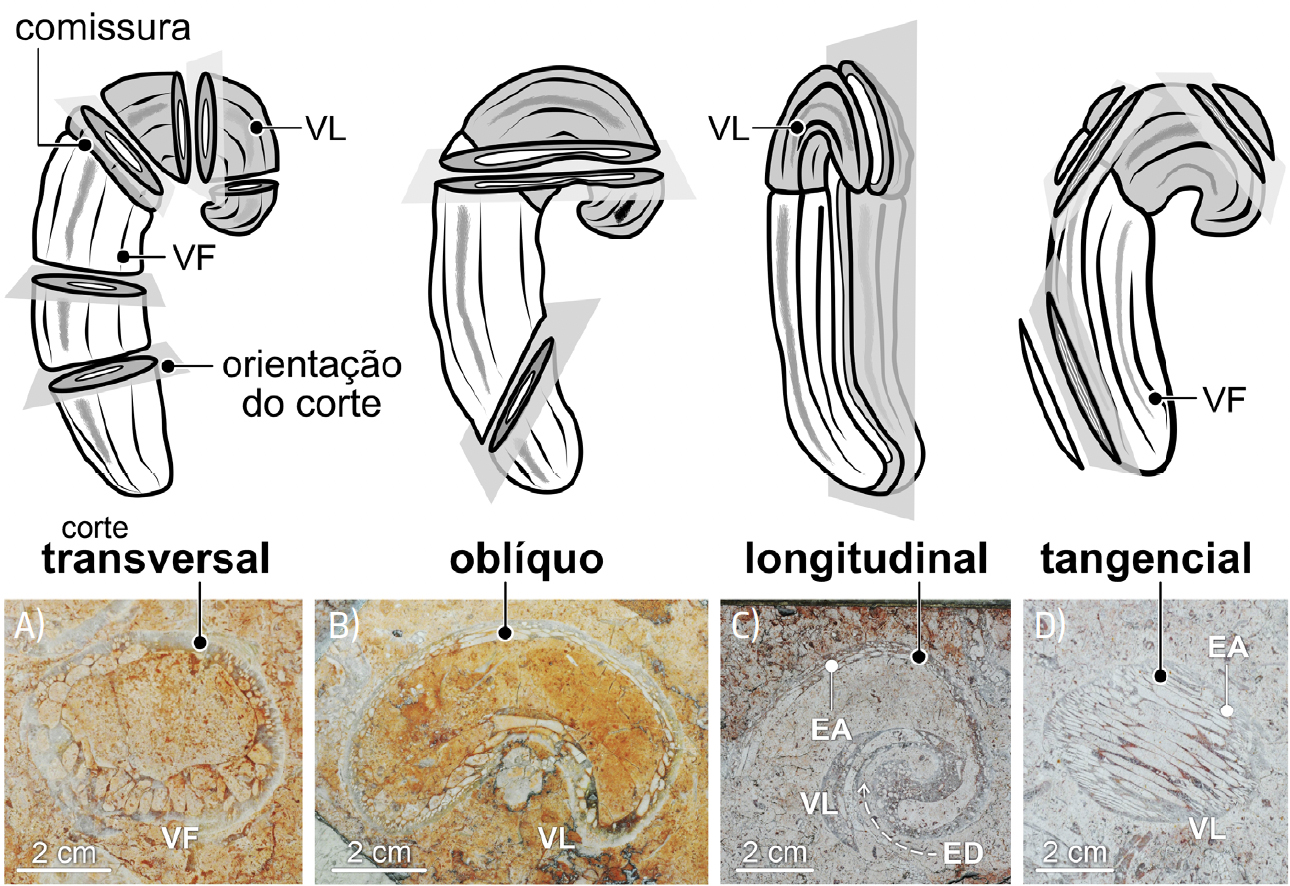
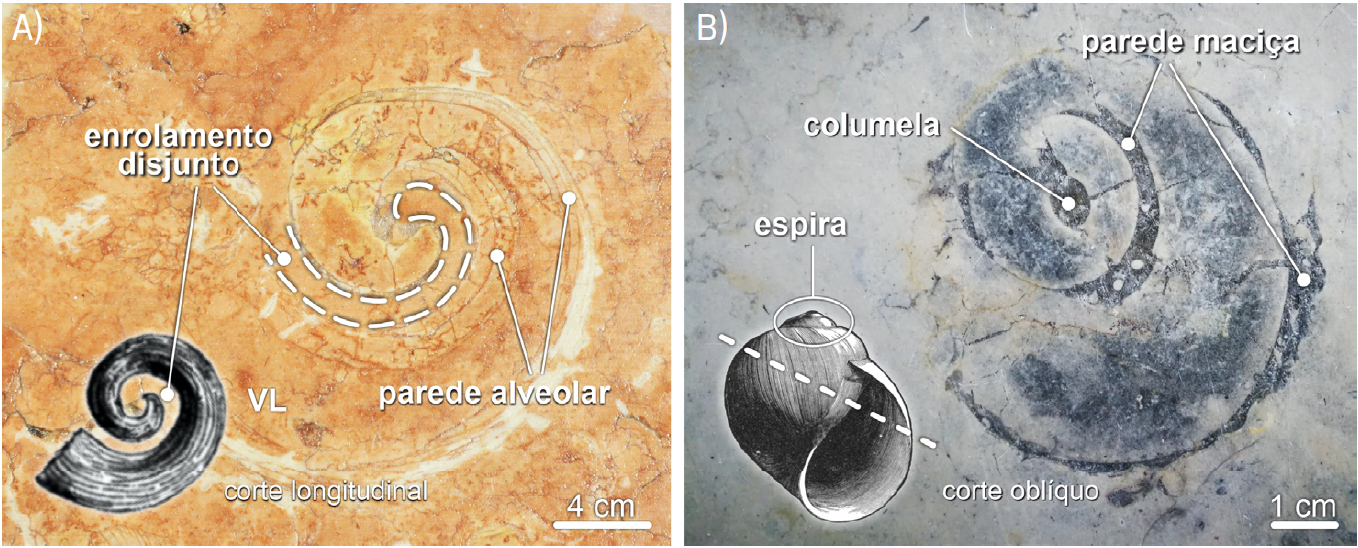
Os fósseis de caprinulídeos, em virtude da forma espiralada da sua valva superior e da estrutura alveolar da parede da concha, são os que apresentam maior diversidade de aspetos em corte (FIGURAS 1B), 14 e 15). De tal modo que cortes longitudinais da valva livre destes rudistas são, por vezes[101], confundidas com secções de conchas de gastrópodes (FIGURA 14C)). Contudo, em secção, os fósseis de caprinulídeos apresentam parede com estrutura alveolar. As valvas livres ostentam, ainda, enrolamento disjunto, i.e., em que as sucessivas voltas da concha não contactam umas com as outras (FIGURA 16), apresentando-se como uma espiral solta, como num corno de cabra (daí o nome Caprinula). As secções oblíquas das valvas livres destes rudistas ostentam, frequentemente, secção reniforme, i.e., em forma de rim (FIGURA 14B)). A comum identificação das secções longitudinais das valvas livres de caprinulídeos como representando fósseis gastrópodes resulta da falsa expectativa de que, por as conchas dos búzios serem espiraladas, os seus fósseis— vistos em corte— também deveriam ser helicoidais. Contudo, consoante a forma da concha dos gastrópodes, os seus cortes transversais podem apresentar morfologias muito variadas, algumas delas nada encaracoladas (FIGURA 17).

Conclusão.
O que fazer com estes fósseis?
As abordagens à geodiversidade urbana são múltiplas, tantas quantos os valores que, em geral, atribuímos às manifestações da natureza geológica. Do puro usufruto estético das rochas ornamentais e dos fósseis que elas encerram, até à sua óbvia vertente económica, passando pelos aspetos culturais e de identidade local, são variadíssimas as maneiras como nos podemos relacionar com os fósseis de rudistas na cidade.
E depois há a vertente lúdica. Do mesmo modo que a observação de aves e de plantas, nomeadamente no contexto citadino em que vivemos, em parques e jardins, promove o bem-estar físico e mental, reduzindo os estados depressivos, a ansiedade e o stress[102], [103], será razoável admitir que a procura, a observação e a identificação de fósseis nesses mesmos contextos, sobretudo em cidades ordenadas, bem mantidas e aprazíveis, trará benefícios similares. A excitação da descoberta de um fóssil nunca antes observado é seguramente similar à da identificação de uma ave ou de uma planta desconhecida. Com a vantagem de que, uma vez descoberto, esse fóssil poderá— se uma deficiente gestão urbana não o obliterar— ser visitado, usufruído e mostrado repetidamente. Não voará dali, migrando para paragens mais aprazíveis no Inverno, nem murchará no final da Primavera.
Na região de Lisboa, por exemplo, o calcário Liós e os seus fósseis de rudistas são parte integrante da identidade da cidade. Estão presentes nos monumentos e nas fachadas de pedra, são representados em pintura de fingimento em painéis de azulejos e em paredes, nas ruas assim como nas nossas casas. São tão característicos de Lisboa como o fado, a sardinha assada e os Santos Populares ou os pastéis de Belém e os Jerónimos. São parte integrante da tessitura da cidade. Sem eles, Lisboa seria um pouco menos Lisboa.
Contudo, talvez a aplicação prática mais óbvia dos fósseis urbanos seja o seu uso no ensino e em atividades de divulgação da Geologia e da geodiversidade. A vertente educativa da geodiversidade em contexto citadino foi já amplamente focada noutras publicações[104], [105], [106] pelo que não será discutida em detalhe aqui. Quer no ensino de temas geológicos, quer biológicos, o potencial dos fósseis urbanos é enorme. Permitem abordar e exemplificar, em concreto, a extinção de grupos biológicos e a sua relação com o uso dos fósseis para investigar a história e a cronologia do planeta. Possibilitam desenvolver em contexto urbano os temas da interpretação paleoambiental e da ligação entre a biodiversidade (aludindo aos organismos do passado) e a geodiversidade (representada pelos seus fósseis). São também um excelente ponto de partida para focar o imperativo ético de proteger a geodiversidade e de conservar a natureza.
É de salientar que uma das muitas vantagens dos fósseis urbanos para o ensino é que para usufruir deles, para os usar em atividades pedagógicas, não é necessário equipamento sofisticado, delicado ou oneroso. Não são imprescindíveis binóculos, nem guias dispendiosos. Também não é necessária roupa especial camuflada. Não há que temer, os fósseis não são assustadiços! Um borrifador com água para os avivar em pavimentos e fachadas, uma câmara fotográfica— ou um qualquer telemóvel— para os registar e curiosidade para os procurar é quanto basta (FIGURA 18).

Apesar de, no nosso país, a publicação de livros de divulgação científica e de manuais de observação da natureza ainda não gozar da popularidade, nem da expressão, que tem noutras paragens, a verdade é que já existem bons guias de observação da geodiversidade urbana em português[107], [108], [109], [110], [111], [112], [113]. Embora tenham sido concebidos tendo em mente a geodiversidade patente nesta ou naquela cidade, o uso destes guias será um excelente ponto de partida para a identificação e o usufruto dos fósseis urbanos em qualquer local.
Dito tudo isto, já só falta uma coisa: agarrar neste ou noutros guias, no borrifador e na câmara fotográfica, sair de casa e começar a procurar, a registar e a usufruir os fósseis que existem mesmo ali ao virar da esquina.
In Memoriam.
Boris Timofeevich Yanin (Борис Тимофеевич Янин, 1931-2021), paleontólogo, estudioso de fósseis de rudistas e saudoso Professor na Cátedra de Paleontologia da Faculdade de Geologia da Universidade Estatal de Moscovo, M.V. Lomonossov. CMS, diplomado em Geologia com especialização em Paleontologia pela UEM em 1986, foi seu discípulo na cadeira de Tafonomia e Paleoecologia, devendo-lhe muito da sua formação paleontológica e recordando-o com saudade.

[editar] Referências
- ↑ SILVA, C. M. & PEREIRA, S., A geodiversidade urbana como recurso educativo, Rev. Ciência Elem., 9, 3. 2021.
- ↑ PEREIRA, H. & LOPES, F., Histórias gravadas nas rochas: Guia de campo: À descoberta das rochas e fósseis em Loulé, Câmara Municipal de Loulé. 2019.
- ↑ RODRIGUES, L. & AGOSTINHO, M., Lagos. Guia de Geologia e de Paleontologia Urbana, Ciência Viva, Câmara Municipal de Lagos. 2016.
- ↑ SILVA, C. M., Geodiversidade no Campus da FCUL, Fósseis de crinóides, Folheto. 2014.
- ↑ SILVA, C. M. & CACHÃO, M., Paleontologia Urbana: Percursos citadinos de interpretação e educação (paleo)ambiental, Comunicações do Instituto Geológico e Mineiro, 84, 2, H33-H35. 1998.
- ↑ SILVA, C. M., Temas de Paleontologia: Somatofóssil, Website. 2006.
- ↑ SILVA, Z., Lioz, rocha património, foi real em Portugal e transportou arte e cultura para o Brasil, Boletim de Minas, 54, 137-146. 2019.
- ↑ SILVA, Z., Lioz - A royal stone in Portugal and a monumental stone in colonial Brazil, Geoheritage, 11, 165-175. 2019.
- ↑ COELHO, C., Estudo preliminar da pedreira romana e outros vestígios identificados no sítio arqueológico de Colaride, Revista Portuguesa de Arqueologia, 5, 2, 277-323. 2002.
- ↑ DEL LAMA, E. A. & COSTA, A. G., Global heritage stones in Brazil, Geoheritage, 14, 25. 2022.
- ↑ SKELTON, P. W., Rudist classification for the revised Bivalvia volumes of the ‘Treatise on Invertebrate Paleontology, Caribbean Journal of Earth Science, 45, 9-33. 2013.
- ↑ NEWELL, N. D., Classification of the Bivalvia, American Museum Novitates, 2206, 1-25. 1965.
- ↑ JOHNSON, C. C., The rise and fall of rudist reefs. Reefs of the dinosaur era were dominated not by corals but by odd mollusks, which died off at the end of the Cretaceous from causes yet to be discovered, American Scientist, 90, 2, 148-153. 2002.
- ↑ SANO, S. I. et al., Late Jurassic-earliest Cretaceous rudists from the Torinosu-type limestones in Southwest Japan, preliminary report, Memoir of the Fukui Prefectural Dinosaur Museum, 7, 67-81. 2008.
- ↑
- ↑ SILVA, C. M., E se os dinossáurios tivessem sido exterminados por um vírus?, Al-Madan, II sér., 23, 8-10. 2020.
- ↑ SILVA, C. M. & COSTA, A. M., Da origem da Terra à ocupação humana: Uma história geológica em Alvalade, Santiago do Cacém, In Memórias da Terra, das Águas e dos Povos, Deus, M. (Coor.), CM Santiago do Cacém, pp. 23-36. 2021.
- ↑ LAMARCK, J. B., Histoire naturelle des animaux sans vertèbres, présentant les caractères généraux et particuliers de ces animaux, 6, 1, 1-343. 1819.
- ↑ LAMARCK, J. B., Histoire naturelle des animaux sans vertèbres, présentant les caractères généraux et particuliers de ces animaux, 6, 1, 1-343. 1819.
- ↑ ORTEGA-HERNÁNDEZ, J., Fossils explained, Rudists, Geology Today, 27, 2, 74-77. 2011.
- ↑ BRASIER, M. D. et al., Changing the picture of Earth’s earliest fossils (3.5–1.9 Ga) with new approaches and new discoveries, Proceedings of the National Academy of Sciences, 112, 16, 4859-4864. 2015.
- ↑ RAUP, D. M., The role of extinction in evolution, PNAS, 91, 15, 6758-6763. 1994.,
- ↑ HULL, P., Life in the aftermath of mass extinctions, Current Biology, 25, 19, R941-R952. 2015.
- ↑ SOLOMON, R., Guggenheim Museum, Form follows function, Website. 2022.
- ↑ MARTINELL, J. et al., Moluscos. In Paleontología de Invertebrados, Martínez Chácon, M.L, Rivas, P., Soc. Española de Paleontología, IGME, Univ. Oviedo, Univ. Granada, 227-376. 2009.
- ↑ POMAR, L. & KENDALL, C., Architecture of carbonate platforms: a response to hydrodynamics and evolving ecology, Controls on carbonate platform and reef development, SEPM Special Publication, 89, 87-216. 2008.
- ↑ POMAR, L. & KENDALL, C., Architecture of carbonate platforms: a response to hydrodynamics and evolving ecology, Controls on carbonate platform and reef development, SEPM Special Publication, 89, 87-216. 2008.
- ↑ POMAR, L., Types of carbonate platforms: a genetic approach, Basin Research, 13, 313-334. 2001.
- ↑ International Chronostratigraphic Chart, International Commission on Stratigraphy. 2022.
- ↑ SHA, J. et al., Paleobiogeographic distribution of rudist bivalves (Hippuritida) in the Oxfordian–early Aptian (Late Jurassic–Early Cretaceous), Cretaceous Research, 108, 104289. 2020.
- ↑ JØRGENSEN, C. B., Bivalve filter feeding revisited, Marine Ecology Progress Series, 142, 287-302. 1996.
- ↑ MARTINELL, J. et al., Moluscos. In Paleontología de Invertebrados, Martínez Chácon, M.L, Rivas, P., Soc. Española de Paleontología, IGME, Univ. Oviedo, Univ. Granada, 227-376. 2009.
- ↑ GILI, E. & SKELTON, P. W., Factors regulating the development of elevator rudist congregations, Geological Society, London, Special Publications, 178, 109-116. 2000.
- ↑ GILI, E. et al., Paleoecology of rudists, Treatise Online, 103, 1–29. 2018.
- ↑ SKELTON, P. W., Constructional morphology and evolution, Springer, Berlin, pp. 375-388. 1991.
- ↑ GILI, E. & SKELTON, P. W., Classificació paleoecològica de les formes dels rudistes - una eina per a l’anàlisi paleoambiental, Butlletí de la Institució Catalana d’Història Natural, 61, 97-116. 1994.
- ↑ GILI, E. & SKELTON, P. W., Factors regulating the development of elevator rudist congregations, Geological Society, London, Special Publications, 178, 109-116. 2000.
- ↑ ORTEGA-HERNÁNDEZ, J., Fossils explained, Rudists, Geology Today, 27, 2, 74-77. 2011.
- ↑ COCITO, S., Bioconstruction and biodiversity: their mutual influence, Scientia Marina, 68, 1, 137-144. 2004.
- ↑ GILI, E. et al., Paleoecology of rudists, Treatise Online, 103, 1–29. 2018.
- ↑ GILI, E. et al., Rudists as gregarious sediment-dwellers, not reef-builders, on Cretaceous carbonate platforms, Palaeogeography, Palaeoclimatology, Palaeoecology, 118, 3-4, 245-267. 1995.
- ↑ SKELTON, P. W., Introduction to the Hippuritida (Rudists): Shell Structure, Anatomy, and Evolution, Treatise Online, 104, 1–37. 2018.
- ↑ GILI, E. et al., Paleoecology of rudists, Treatise Online, 103, 1–29. 2018.
- ↑ GILI, E. et al., Rudists as gregarious sediment-dwellers, not reef-builders, on Cretaceous carbonate platforms, Palaeogeography, Palaeoclimatology, Palaeoecology, 118, 3-4, 245-267. 1995.
- ↑ GILI, E. & SKELTON, P. W., Factors regulating the development of elevator rudist congregations, Geological Society, London, Special Publications, 178, 109-116. 2000.
- ↑ MASSE, J.-P. & PHILIP, J., European Fossil Reef Models, Society of Economic Paleontologists and Mineralogists, 30, 399-426. 1981.
- ↑ SKELTON, P. W. et al., Rudists as successful sediment-dwellers, not reef-builders, on Cretaceous carbonate platforms, The Paleontological Society Special Publications, 6, 271-271. 1992.
- ↑ WOOD, R., The ecological evolution of reefs, Annual Review of Ecology and Systematics, 29, 1, 179-206. 1998.
- ↑ WOOD, R., Taphonomy of reefs through time. In: Taphonomy, Springer, Dordrecht, pp. 375-409, 2011.
- ↑ RIDING, R., Structure and composition of organic reefs and carbonate mud mounds: concepts and categories, Earth-Science Reviews, 58, 1-2, 163-231. 2002.
- ↑ RIDING, R., Structure and composition of organic reefs and carbonate mud mounds: concepts and categories, Earth-Science Reviews, 58, 1-2, 163-231. 2002.
- ↑ SCHAFHAUSER, A. et al., Depositional Environment of coral-rudist associations in the Upper Cretaceous Cardenas Formation (Central Mexico), Geologia Croatica, 56, 2, 187-198. 2003.
- ↑ GILI, E. & SKELTON, P. W., Factors regulating the development of elevator rudist congregations, Geological Society, London, Special Publications, 178, 109-116. 2000.
- ↑ GÖTZ, S., Biotic interaction and synecology in a Late Cretaceous coral–rudist biostrome of southeastern Spain, Palaeogeography, Palaeoclimatology, Palaeoecology, 193, 1-14. 2003.
- ↑ CALLAPEZ, P. M., Palaeogeographic evolution and marine faunas of the mid-Cretaceous Western Portuguese carbonate platform, Thalassas, 24, 1, 29-52. 2008.
- ↑ HATTORI, K. et al., Sequence stratigraphic and paleoecologic analysis of an Albian coral-rudist patch reef, Arizona, USA, Palaios, 34, 12, 600-615. 2019.
- ↑ MASSE, J. P. & PHILIP, J., European Fossil Reef Models, SEPM (Society for Sedimentary Geology) Special Publication, 30, 399–426. 1981.
- ↑ GILI, E. et al., Corals to rudists—an environmentally induced assemblage succession, Palaeogeography, Palaeoclimatology, Palaeoecology, 119, 1–2, 127-136. 1995.
- ↑ PACHECO, M. & CACHÃO, M., Urban geology of Lisbon: The importance of the National Palace of Ajuda (Lisbon, Portugal), Geoheritage, 13, 3, 1-13. 2021.
- ↑ HARTSHORNE, P. M., Facies architecture of a Lower Cretaceous coral-rudist patch reef, Arizona, Cretaceous Research, 10, 4, 311-336. 1989.
- ↑ WILSON, J. L., Carbonate facies in geologic history, Springer Verlag, Nova Iorque, 471 pp. 1975.
- ↑ MASSE, J. P. & PHILIP, J., European Fossil Reef Models, SEPM (Society for Sedimentary Geology) Special Publication, 30, 399–426. 1981.
- ↑ ORTEGA-HERNÁNDEZ, J., Fossils explained, Rudists, Geology Today, 27, 2, 74-77. 2011.
- ↑ GILI, E. et al., Rudists as gregarious sediment-dwellers, not reef-builders, on Cretaceous carbonate platforms, Palaeogeography, Palaeoclimatology, Palaeoecology, 118, 3-4, 245-267. 1995.
- ↑ SKELTON, P. W. & GILI, E., Rudists and carbonate platforms in the Aptian: a case study on biotic interactions with ocean chemistry and climate, Sedimentology, 59, 1, 81-117. 2012.
- ↑ ROSS, D. J. & SKELTON, P. W., Rudist formations of the Cretaceous: a palaeoecological, sedimentological and stratigraphical review, Sedimentology Review, 1, 73-91. 1993.
- ↑ MARTINELL, J. et al., Moluscos. In Paleontología de Invertebrados, Martínez Chácon, M.L, Rivas, P., Soc. Española de Paleontología, IGME, Univ. Oviedo, Univ. Granada, 227-376. 2009.
- ↑ GILI, E. et al., Paleoecology of rudists, Treatise Online, 103, 1–29. 2018.
- ↑ GILI, E. et al., Rudists as gregarious sediment-dwellers, not reef-builders, on Cretaceous carbonate platforms, Palaeogeography, Palaeoclimatology, Palaeoecology, 118, 3-4, 245-267. 1995.
- ↑ GILI, E. et al., Rudists as gregarious sediment-dwellers, not reef-builders, on Cretaceous carbonate platforms, Palaeogeography, Palaeoclimatology, Palaeoecology, 118, 3-4, 245-267. 1995.
- ↑ SKELTON, P. W. & GILI, E., Rudists and carbonate platforms in the Aptian: a case study on biotic interactions with ocean chemistry and climate, Sedimentology, 59, 1, 81-117. 2012.
- ↑ SILVA, C. M., Temas de Paleontologia: Icnofóssil, Website. 2006.
- ↑ FOSSIL ID, Thalassinoides, Geoscience Data and Collections. 2020.
- ↑ FOSSIL ID, Thalassinoides, Geoscience Data and Collections. 2020.
- ↑ Official State Fossils, National Parks Service. Fossils and Paleontology.
- ↑ List of State Fossils, Fossilera.
- ↑ SILVA, C. M., Geodiversidade no Campus da FCUL, Estilólitos, Website. 2014.
- ↑ ORNABASE, Rochas ornamentais portuguesas, LNEG. 2012.
- ↑ SOARES, C. M. et al., As rochas ornamentais na Basílica de Nossa Senhora do Rosário de Fátima (Portugal): entre o estético e o simbólico, BSAA Arte, 88, 371-399. 2022.
- ↑ ORNABASE, Rochas ornamentais portuguesas, LNEG. 2012.
- ↑ COELHO, C., Estudo preliminar da pedreira romana e outros vestígios identificados no sítio arqueológico de Colaride, Revista Portuguesa de Arqueologia, 5, 2, 277-323. 2002.
- ↑ SILVA, Z., Lioz, rocha património, foi real em Portugal e transportou arte e cultura para o Brasil, Boletim de Minas, 54, 137-146. 2019.
- ↑ SILVA, Z., Lioz - A royal stone in Portugal and a monumental stone in colonial Brazil, Geoheritage, 11, 165-175. 2019.
- ↑ DEL LAMA, E. A. & COSTA, A. G., Global heritage stones in Brazil, Geoheritage, 14, 25. 2022.
- ↑ CANELAS, L., Há um padrão português do séc. XV encaixotado desde 2019 num porto na Namíbia, Público, Ano 34, Nº 12011. 2023.
- ↑ SILVA, C. M. & PEREIRA, S., Walking on geodiversity: the artistic stone-paved sidewalks of Lisbon (Portugal) and their heritage value, Geoheritage, 14, 98. 2022.
- ↑ UNIVERSIDADE ABERTA, Motivos geológicos nos azulejos do Palácio Ceia, Vídeo educativo sobre os motivos geológicos nos azulejos do Palácio Ceia, com depoimentos de Alexandra Câmara e Carlos Marques da Silva. 2019.
- ↑ SILVA, C. M., Urban geodiversity and decorative arts: the curious case of the “Rudist Tiles” of Lisbon (Portugal), Geoheritage, 11, 151-163.
- ↑ BANDEL, K. & MUSTAFA, H., Constructional morphology of some Upper Cretaceous rudists of the Ajlun (Jordan), Mitteilungen aus dem Geologisch-Paläontologischen Institute der Universität Hamburg, 77, 603-635. 1996.
- ↑ MARTINELL, J. et al., Moluscos. In Paleontología de Invertebrados, Martínez Chácon, M.L, Rivas, P., Soc. Española de Paleontología, IGME, Univ. Oviedo, Univ. Granada, 227-376. 2009.
- ↑ GILI, E. et al., Paleoecology of rudists, Treatise Online, 103, 1–29. 2018.
- ↑ SHARPE, D., On the secondary district of Portugal which lies on the north of the Tagus, Quarterly Journal of the Geological Society, 6, 1-2, 135-201. 1850.
- ↑ DOUVILLÉ, H., Études sur les caprines, Bulletin de la Société Géologique de France, 3, 16, 699-730. 1888.
- ↑ DECHASEAUX, C. et al., Treatise on Invertebrate Paleontology, Part N, Mollusca 6, Bivalvia, 2, University of Kansas, Lawrence, Kansas and Geological Society of America, N749-817. 1969.
- ↑ SKELTON, P. W., Rudist classification for the revised Bivalvia volumes of the ‘Treatise on Invertebrate Paleontology, Caribbean Journal of Earth Science, 45, 9-33. 2013.
- ↑ SKELTON, P. W. & SMITH, A. B., The evolutionary biology of the Bivalvia, Geological Society, London, Special Publications, 177, 97-127.
- ↑ YANIN, B.T., Sistematika i Filogeniia Bespozvonochikh. Kriterii Vydeleniia Vyshikh Taksonov, Academy of Sciences of the USSR, Moscow Society of Naturalists, Moscovo, pp. 57-69. 1990.
- ↑ YANIN, B. T., The system, phylogeny and evolution of rudists, Palaeontological Institute of the Russian Academy of Sciences, Moscovo, 228 pp. 1995.
- ↑ DECHASEAUX, C. et al., Treatise on Invertebrate Paleontology, Part N, Mollusca 6, Bivalvia, 2, University of Kansas, Lawrence, Kansas and Geological Society of America, N749-817. 1969.
- ↑ DECHASEAUX, C. et al., Treatise on Invertebrate Paleontology, Part N, Mollusca 6, Bivalvia, 2, University of Kansas, Lawrence, Kansas and Geological Society of America, N749-817. 1969.
- ↑ MOZER, A. et al., Mapping Lioz limestone in monuments at Rio de Janeiro, Brazil, Geoheritage, 14, 2, 1-14. 2022.
- ↑ UNIVERSITY OF EXETER, Watching birds near your home is good for your mental health: People living in neighborhoods with more birds, shrubs and trees are less likely to suffer from depression, anxiety and stress, ScienceDaily. 2017.
- ↑ MASSE, J. P. & PHILIP, J., European Fossil Reef Models, SEPM (Society for Sedimentary Geology) Special Publication, 30, 399–426. 1981.
- ↑ SILVA, C. M. & PEREIRA, S., A geodiversidade urbana como recurso educativo, Rev. Ciência Elem., 9, 3. 2021.
- ↑ SILVA, C. M., Geoscience Education Indoor and Outdoor, Science Education, Springer Verlag, pp. 133-156. 2016.
- ↑ PEREIRA. H. & LOPES, F., Geodiversity in our school’s walls and pavements, IV Encontro Internacional Casa das Ciências, Livro de Resumos, pp. 89-90. 2017.
- ↑ SILVA, C. M. & PEREIRA, S., A geodiversidade urbana como recurso educativo, Rev. Ciência Elem., 9, 3. 2021.
- ↑ PEREIRA, H. & LOPES, F., Histórias gravadas nas rochas: Guia de campo: À descoberta das rochas e fósseis em Loulé, Câmara Municipal de Loulé. 2019.
- ↑ RODRIGUES, L. & AGOSTINHO, M., Lagos. Guia de Geologia e de Paleontologia Urbana, Ciência Viva, Câmara Municipal de Lagos. 2016.
- ↑ SILVA, C. M., Geodiversidade no Campus da FCUL, Fósseis de crinóides, Folheto. 2014.
- ↑ RODRIGUES, L. & AGOSTINHO, M., Faro. Guia de Geologia e de Paleontologia Urbana, Ciência Viva, Câmara Municipal de Lagos. 2016.
- ↑ RODRIGUES, L. & AGOSTINHO, M., Tavira. Guia de Geologia e de Paleontologia Urbana, Ciência Viva, Câmara Municipal de Lagos. 2016.
- ↑ PEREIRA, H. & LOPES, F., Histórias gravadas nas rochas, Guia de campo: À descoberta das rochas e fósseis em Loulé, Câmara Municipal de Loulé. 2019.
Criada em 23 de Maio de 2023
Revista em 13 de Junho de 2023
Aceite pelo editor em 15 de Dezembro de 2023
