Mutações
Referência : Amorim, A., (2023) Mutações, Rev. Ciência Elem., V11(3):030
Autora: António Amorim
Editor: João Nuno Tavares
DOI: [https://doi.org/10.24927/rce2023.030]

[editar] Resumo
As mutações são raras e a estimativa da frequência com que ocorrem é difícil. Ocorrem em muito diversas formas e as consequências são correspondentemente variadas. Analisam-se aqui esses dois problemas na perspetiva evolutiva.
Mutação (mudança, no nosso latim vulgar) não é mais que um erro de cópia (ou o seu resultado) do material genético (só abordaremos o que tecnicamente se designa por mutação germinal, isto é, as mutações que ocorrem na produção de gâmetas que originem um descendente, por oposição às somáticas, que ocorrem nas células não reprodutoras). Este erro, “gato” ou “gralha” na imagem tipográfica, pode assumir muitas formas e implicar consequências muito diversas e é o motor da evolução. Adivinha-se, portanto, que a velocidade evolutiva será dependente da frequência com que estes erros ocorrerão. Também será fácil inferir que em muitos casos as implicações para a reprodução do organismo portador da mutação poderão ser tão graves que impeçam a sua reprodução e nesse caso o erro será literalmente um nado- morto.
Tipos de mutação.
Vamos então começar por dar uma ideia dos tipos de mutação que podem ocorrer para podermos concretizar estas intuições. Para isso teremos de lembrar que a informação genética está contida na sequência e organização de um ácido nucleico em todos os atuais seres vivos conhecidos (DNA, ou RNA em alguns vírus). A codificação da informação é feita através da variação da composição química do DNA em termos dos seus nucleótidos, simbolizados pelas letras A, C, G e T. Uma sequência de DNA poderá, pois, ser representada como um texto de um alfabeto reduzido, por exemplo:
…ATCGATTCAGA…
Um dos possíveis erros será uma troca de letras, uma substituição:
…ATCGATTCACA…
em que o original G foi erradamente copiado e introduzido um C, sublinhado na sequência mutada. Note-se que, apesar de se tratar de uma letrinha apenas, as consequências podem ser dramáticas e levar à incapacidade de sobrevivência do seu portador, o que significa que a seriedade das consequências não é previsível pela simples análise do erro, fora do seu contexto. Por outras palavras, uma mesma substituição (no nosso caso, G→C) pode ser absolutamente inócua ou letal, dependendo da leitura dessa informação.
Note-se que as substituições não acarretam variação do comprimento da sequência; pelo contrário, uma deleção ou uma inserção implicarão diminuição ou aumento, respetivamente.
Temos ainda um conjunto de mutações em que se alteram a ordem ou a posição de sequências, como translocações (literalmente, mudança de local) e inversões. Uma vez que o material genético nos seres vivos complexos está organizado em várias unidades, arcaicamente chamadas cromossomas (como livros numa biblioteca) teremos ainda a possibilidade de observar mudanças na arquitetura do material genético, correspondentes a variação do seu número (nulissomias, monossomias, trissomias, etc.) ou da sua estrutura (inversões, duplicações), podendo ainda ocorrer fusões cromossómicas.
Tal como dissemos a propósito das substituições, todas estas alterações mutacionais tem um grau muito variável de consequências para os seus portadores, mas algumas interferem sempre com a sua capacidade reprodutiva e, portanto, são especialmente importantes na evolução. Veremos isso no final deste trabalho, mas antes tentaremos responder à questão já levantada, da frequência das mutações.
Deteção das mutações e estimativa da sua frequência.
Como dissemos, algumas mutações acarretam consequências dramáticas para os seus portadores. Nesses casos, que eram os acessíveis antes da vulgarização das análises bioquímicas e moleculares, a sua ocorrência era imediatamente detetada. Um dos exemplos mais flagrante e suficientemente comum para todos termos já observado um caso, era o do nanismo acondroplásico: pais com estatura normal geravam um filho com esta anomalia, apenas explicável por mutação. Para além desta forma de deteção, que podemos chamar genealógica, existe uma outra, indireta, a filogenética, em que a ocorrência de mutação é inferida do facto de duas espécies com um ancestral comum terem informação genética distinta. Muito simplificada e resumidamente, se observamos que Homo sapiens tem, em uma localização específica do genoma, a sequência:
ATCGATTCAGA
e Pan troglodytes na posição homóloga:
ATCGATTCACA
inferimos que algures na evolução até estas espécies atuais, terá ocorrido uma mutação. Note-se que dispondo apenas desta informação não sabemos qual o estado original, isto é, se a mutação terá sido G→C, C→G, ou ainda se o estado no ancestral comum às duas espécies era outro, A ou T. Nesta abordagem fica, consequentemente, em aberto a possibilidade de ter ocorrido mais do que uma mutação, assim como não se pode descartar a hipótese da ocorrência de mutações sucessivas reversas, que restaurem o estado ancestral, por exemplo C→G seguida de G→C.
Dada a complexidade deste método de deteção e da cronologia associada, vamos voltar ao outro tipo de abordagem, a genealógica. Atualmente existem meios técnicos que nos permitem, com razoável fiabilidade, determinar a sequência da maioria do material genético de um organismo. Esta possibilidade levar-nos-ia a pensar que poderíamos detetar todas as mutações desde que sequenciássemos ambos os progenitores e um seu descendente. Tal não é, porém, o caso: mesmo com o recurso às tecnologias mais modernas, uma fração importante das mutações não é detetada. Vamos demonstrá-lo recorrendo a um exemplo que tem igualmente relevância forense, isto é que põe problemas na interpretação de resultados destinados a validar um parentesco genético, como o de paternidade ou maternidade.
Suponhamos que estudamos uma região do genoma cujo modo de transmissão é diploide homogamético (o tratado pela versão original da teoria mendeliana), ou seja, em que cada indivíduo tem duas cópias da informação genética, mas só transmite à descendência, de forma aleatória, uma delas:
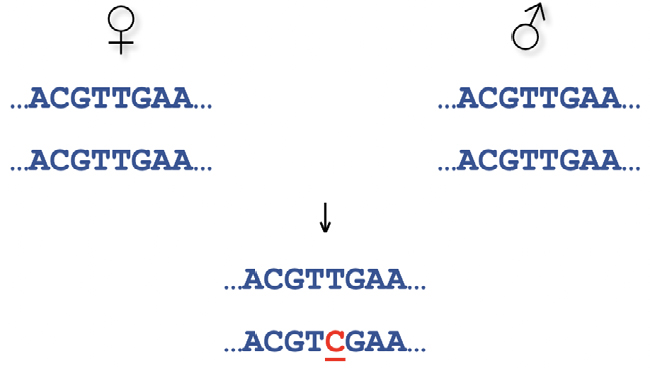
Claramente, o descendente evidencia uma mutação, inferida da incompatibilidade mendeliana: na 5.ª posição da sequência aparece C, quando ambos os progenitores são T, o que significa que em um deles (não sabemos se o masculino ou o feminino), terá havido um erro de cópia, uma substituição. Repare-se que neste exemplo assumimos como verdadeiros ambos os parentescos, mas podemos enfrentar uma situação em que esteja em dúvida a paternidade. Nesse caso seria possível interpretar a mesma observação como resultante da inexistência desse parentesco e não de uma mutação. Ora não será possível quantificar a plausibilidade relativa destas duas interpretações alternativas sem termos uma estimativa fiável da frequência desta mutação para a contrapormos à probabilidade de C ser a contribuição de um desconhecido (o verdadeiro pai).
Pelo que vimos anteriormente, seria simples: bastaria contar o número de incompatibilidades mendelianas observadas numa amostra de trios pai/mãe/filho, como a esquematizada na tabela seguinte, onde as informações genéticas alternativas são simbolizadas por 1 e 2 e se assinalam a amarelo as células onde se verificam incompatibilidades mendelianas e, por conseguinte, se terá verificado mutação:
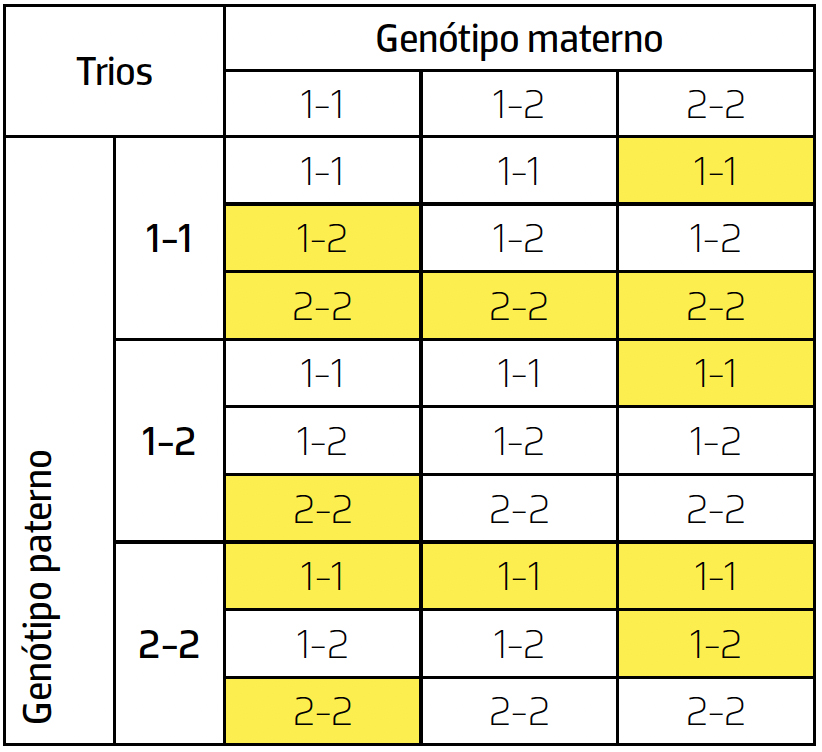
A estimativa da frequência das mutações (vulgarmente designada por taxa de mutação) nesta amostra será, pois, obtida simplesmente pelo somatório das frequências das células a amarelo (Na verdade, embora este seja o processo comum de proceder a esta estimativa, devemos esclarecer que é incorreto; em algumas células (por exemplo na correspondente ao trio 1-1/1-1/2-2, terão corrido duas mutações), bem como o total de observações a considerar não é o total de trios contabilizado, mas sim o seu dobro, dado que em cada trio observamos dois eventos de cópia, paterna e materna).
Esta estimativa é, no entanto, enviesada: vamos finalmente abordar a questão sugerida no título: as mutações escondidas, que serão essenciais para uma estimativa correta da frequência das mutações.
Para isso suponhamos agora uma situação em que não se verifique uma incompatibilidade mendeliana:
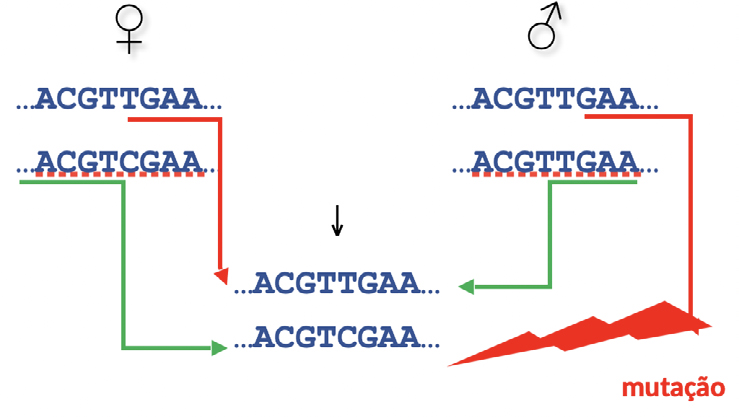
Neste caso, a interpretação mais tentadora, que não invoca mutação, é a de que a sequência com C na 5.ª posição da sequência do descendente tenha sido transmitida pelo verdadeiro pai, como ilustrado pelas setas verdes. Existe, no entanto, uma outra, como evidenciado pelas setas vermelhas, que demonstra a possibilidade da ocorrência (escondida) de mutação.
Mas esta possibilidade será quantitativamente relevante, ou seja, a proporção de mutações escondidas será significativa? Sem entrar em pormenores tecnicamente complexos, podemos afirmar que sim; olhemos para a tabela seguinte, onde se apresentam os possíveis descendentes em termos de serem (M) ou não (N) resultado de mutação (de novo, para simplificar a apresentação, as duas composições alternativas das sequências analisadas são representadas por 1 e 2).
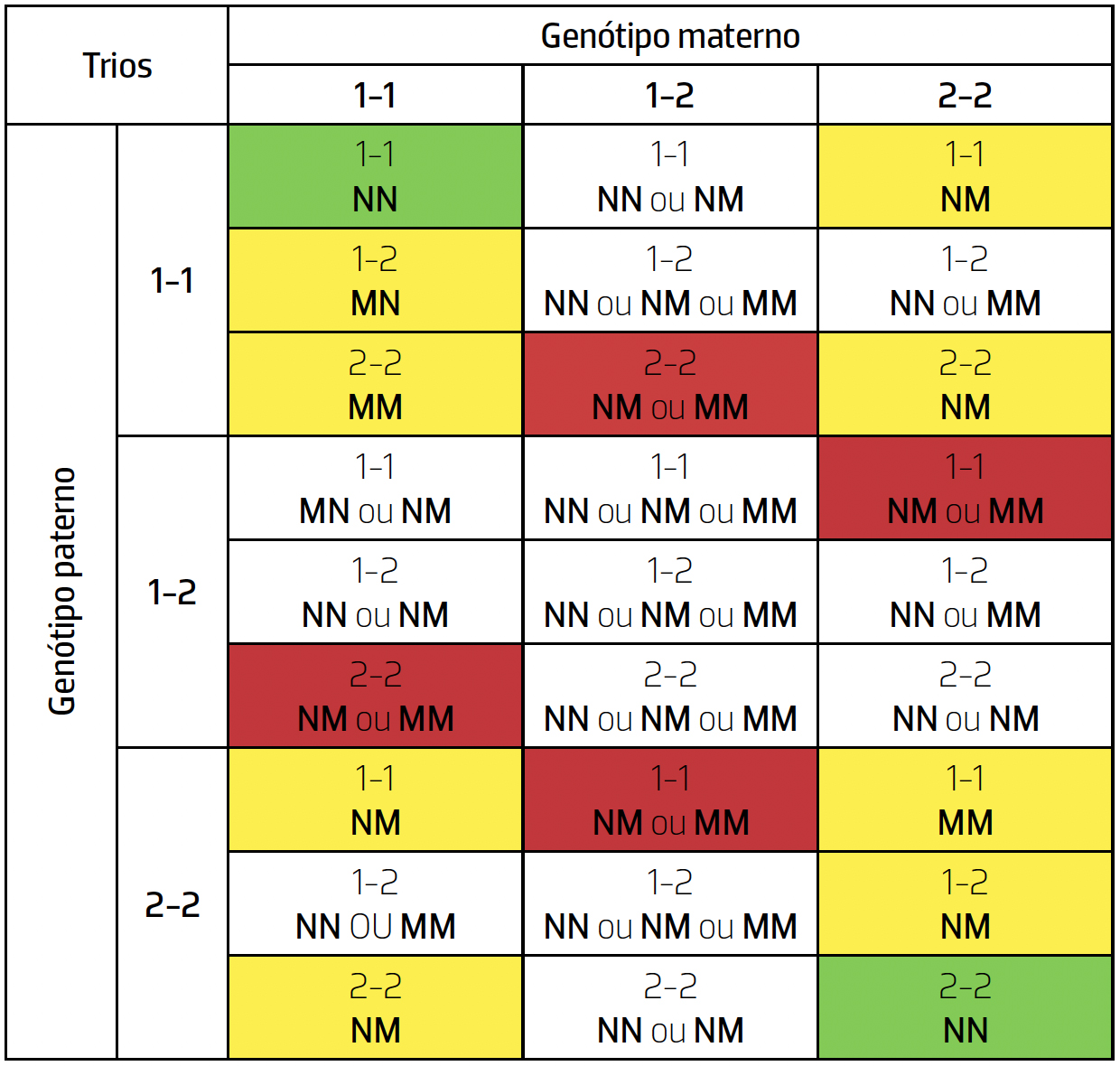
Vemos que a consideração da possibilidade de existirem mutações que não impliquem incompatibilidades mendelianas introduz complexidades inesperadas: a origem de vários trios torna-se ambígua. Por exemplo: o trio 1-1/1-2/1-1 pode resultar da ausência de mutação (tanto paterna como materna), mas também da ausência de mutação paterna combinada com mutação materna e o trio 1-1/1-2/1-2 tem como possível origem a ausência de mutação em ambos os progenitores, mutação em apenas um deles e ainda mutação em ambos. Em resumo:
- Apenas nas células verdes podemos garantir que não houve mutação;
- Nas células amarelas podemos garantir que houve mutação e o seu número (em alguns trios uma e em outros duas);
- Nas células vermelhas podemos garantir que houve mutação, mas não o seu número (o mesmo trio pode resultar de uma ou duas mutações);
- As células não coloridas podem ocorrer com ou sem mutação.
Ora são estas últimas, correspondentes a casos em que não se verifica incompatibilidade mendeliana, que podem então “esconder” mutações. Não tem aqui lugar a derivação analítica da sua frequência esperada, mas fica claro que a metodologia corrente (contagem de incompatibilidades mendelianas) para determinar taxas de mutação subestimará o seu verdadeiro valor.
Mutações e evolução.
Afirmou-se no início que as mutações são o motor da evolução. Neste ponto, em que “descobrimos” ser a taxa de mutação subestimada, estaríamos pelo menos tentados a concluir que então a evolução deveria ser mais rápida que a prevista. Esta intuição é porém falaciosa por duas ordens de razões: não definimos o que entendemos por “evolução” e a definição de taxa de mutação que usámos omite o caráter direcional das mutações.
Comecemos pela primeira: evolução, tanto em genética como no sentido comum, significa mudança, mas em genética, a mudança reveste-se de duas formas biologicamente muito distintas nos seres vivos complexos com reprodução sexuada: a (micro)evolução que aborda as mudanças de frequência das características genéticas no seio de uma espécie ao longo do tempo e a (macro)evolução, que se ocupa do aparecimento de novas espécies. Tudo o que tratámos até agora respeita apenas a microevolução, e o papel das mutações na macroevolução será deixado para a última secção.
Quanto à questão da direccionalidade mutacional, relembremos que no modelo simples que utilizámos, que se aplica apenas a um tipo de mutação (substituição), supusemos um sítio do genoma, onde, na população em estudo coexistiam duas versões (sem impacto diferencial no potencial reprodutivo) na ocupação de um local genómico (que simbolizámos por 1 e 2) e indicámos como se calcularia não enviesadamente a frequência global dos erros de transmissão (1→2 e 2→1). Ora este parâmetro (taxa de mutação por sítio do genoma) nada nos diz de útil relativamente à previsão da evolução das frequências de 1 e de 2. De facto, qualquer que seja a taxa média, se a frequência da mutação 1→2 for maior que a simétrica, 2→1, o sistema evoluirá, na ausência de outros fatores, para uma situação em que as frequências de 1 e 2 serão proporcionais às taxas de mutação; no caso limite, se a taxa de mutação 2→1 for igual a 0, verificar-se-á a fixação da forma 1 e toda a população ficará geneticamente uniforme quanto a este local do genoma.
Assim, paradoxalmente, um local com uma alta taxa de mutação pode não evoluir (se as mutações simétricas forem equifrequentes), e em um outro, ainda que com baixa taxa de mutação, pode observar-se evolução (se a mutação for unidirecional).
Mutações e origem das espécies.
Para abordar a (macro)evolução será necessário definir “espécie”. O conceito só faz realmente sentido quando aplicado a seres vivos com reprodução sexuada, e indica o conjunto de indivíduos que podem potencialmente reproduzir-se entre si gerando descendentes férteis (e incapazes de o fazer com sucesso, ou gerando híbridos inférteis, com membros de outras espécies).
Já abordámos o papel das mutações na evolução intraespecífica. O que nos falta tratar será responder à pergunta: podem as mutações criar o isolamento reprodutor, condição necessária para a formação de espécies? A resposta é afirmativa, e podemos mesmo dizer que sem a ocorrência dessas mutações é difícil conceber a estabilidade do isolamento reprodutor e, portanto, das espécies. Que mutações são essas? Pois são as que envolvem rearranjos do genoma tais que perturbam ou mesmo impedem totalmente a formação de gâmetas reprodutivamente viáveis (as já referidas inversões e fusões cromossómicas, por exemplo). A base biológica para esta disrupção da reprodução está dependente da existência de um mecanismo de verificação e redistribuição da informação genética dos progenitores (a recombinação meiótica) que exige homologia (quase) perfeita entre essas duas versões. Não vamos aqui abordar estes processos nem as formas pelos quais, na célula mãe de gâmetas, a coexistência de versões estruturalmente diferentes do genoma tem estre efeito abortivo.
De facto, quaisquer que sejam os mecanismos biológicos envolvidos, o que podemos concluir é que:
a) Existem mutações que podendo não ter qualquer efeito fisiológico visível, são causadoras de isolamento reprodutor, potencialmente conducentes a especiação e que;
b) As mutações são um duplo motor híbrido da evolução: por um lado, alterando a composição genética das espécies e por outro criando as condições para a formação de novas espécies;
c) A determinação da frequência com que ocorrem as mutações é difícil e tem sido teoricamente mal formulada e sistematicamente enviesada e/ou subestimada.
As confusões conceptuais que têm perdurado na área da biologia evolutiva e que aqui pretendemos contribuir para dissipar, são responsáveis por muitas das erróneas, ainda que populares, visões da evolução. A mais básica e grave será a que consiste em assumir a continuidade dos mecanismos micro e macroevolutivos, pretendendo que especiação não seria mais do que o resultado da acumulação de diferenças que acabariam por levar ao isolamento reprodutor. O que se verifica experimentalmente e aqui tentámos explicitar é o contrário: que (para organismos com reprodução sexuada) o isolamento reprodutor precede a diferenciação.
[editar] Referências
- AMORIM, A., A Espécie das Origens, Gradiva, Lisboa. 2ª ed.. 2009.
- AMORIM, A., A origem das espécies. In BERTOLAMI O, COUTO H (eds) Do Big Bang ao Homem, U. Porto Edições, Porto. 2016.
- AMORIM, A., Mutation rate (under)estimation through Mendelian incompatibilities, Human Population Genetics and Genomics. 2023.
- AMORIM, A. & PINTO, N., Estimates of mutation rates from incompatibilities are misleading - guidelines for publication and retrieval of mutation data urgently needed, Forensic Science International: Genetics Supplement Series, 7, 612-613. 2019.
- LYNCH, M., The Origins of Genome Architecture, Sinauer Associates, Inc. Publishers, Sunderland, MA. 2007.
- ANTÃO-SOUSA, S. et al., Underestimation and misclassification of mutations at X chromosome STRs depend on population’s allelic profile, Forensic Science International: Genetics Supplement Series, 7, 718–720. 2019.
- PINTO N, A. et al., Theory and statistics of mutation rates: A mathematical framework reformulation for forensic applications, Forensic Science International: Genetics Supplement Series, 5, 131–132. 2015.
- PINTO, N. et al., Mutation and mutation rates at Y chromosome specific Short Tandem Repeat Polymorphisms (STRs): A reappraisal, Forensic Science International: Genetics, 9, 20-24. 2014.
Criada em 5 de Junho de 2023
Revista em 3 de Julho de 2023
Aceite pelo editor em 13 de Outubro de 2023
