Big data
Referência : Ávila, S. P., Melo, C., Madeira, P., (2022) Big data, Rev. Ciência Elem., V10(4):051
Autores: Sérgio P. Ávila, Carlos Melo e Patrícia Madeira
Editor: João Nuno Tavares
DOI: [https://doi.org/10.24927/rce2022.051]

Resumo
Para quem, como os autores deste texto, trabalha em ilhas oceânicas e tenta decifrar os processos e padrões evolutivos que aqui decorrem atualmente, bem como os que decorreram ao longo do tempo geológico, assume particular relevo o conhecimento que é necessário ter acerca da ontogenia de cada ilha, ou seja, da história evolutiva de cada uma das ilhas estudadas, bem como da história de cada uma das espécies que, ao longo dos milhões de anos de tempo útil de uma ilha, foi capaz de lá chegar, aí viver e, pelo menos algumas, evoluir, originando espécies endémicas[1]. A grande maioria das ilhas oceânicas passa por vários estados ontogenéticos, com uma história evolutiva que se pode resumir da seguinte forma: 1) fase de monte submarino; 2) fase de ilha emergente; 3) fase de construção de edifícios vulcânicos; 4) fase erosiva; 5) fase de atol (dependente da ocorrência de corais); 6) fase de guyot[2]. Nas fases 1 a 3 predomina a construção da ilha, com altas taxas de emissão de produtos vulcânicos, ao passo que a partir do final da fase 3 predominam os processos erosivos que atuam a par dos usuais processos de subsidência do(s) edifício(s) vulcânico(s), fazendo com que, progressivamente, a ilha se transforme num monte submarino de topo aplanado, conhecido pelo nome de guyot.
Esta descrição é importante pois o impacto, sobre os seres vivos, da história evolutiva de uma ilha oceânica vulcânica é diferente, consoante trabalhemos com organismos marinhos ou com organismos terrestres. É fácil de entender que os organismos marinhos podem colonizar uma proto-ilha quando esta ainda está na fase de monte submarino, ao passo que, somente quando há área emersa, é que há substrato disponível para ser colonizado por organismos terrestres. Por outro lado, quando uma antiga ilha é arrasada por agentes erosivos e passa à fase de monte submarino, os organismos terrestres são extirpados, por vezes milhares de anos antes do desaparecimento dos organismos marinhos litorais, os quais só desaparecem quando, no processo de subsidência, os picos mais elevados do monte submarino descem abaixo de cerca de 50 metros[3].
Para além do conhecimento geológico detalhado que é proporcionado pelas diferentes histórias evolutivas das ilhas, ilhéus e montes submarinos que formam um determinado arquipélago, é importante para os biólogos marinhos darem resposta às seguintes questões: 1) De onde vieram os colonizadores iniciais que povoaram a primeira ilha/monte submarino desse arquipélago? 2) Quando atingiram essa primeira ilha pela primeira vez? 3) O que lhes sucedeu ao longo do tempo evolutivo? Todas estas questões estão atualmente enquadradas por um ramo fascinante da Biologia que dá pelo nome de Biogeografia Insular. Tendo por base os estudos iniciais de Wallace, em 1880<[4], a Biogeografia Insular sofreu um enorme incremento com a obra seminal de MacArthur & Wilson de 1967[5], e teve recentemente desenvolvimentos extraordinários para os investigadores que trabalham com organismos insulares terrestres[6], [7], [8], <[9], [10], bem como para aqueles que estudam organismos insulares marinhos[11], [12], [13], [14], [15].
Enquanto que o estudo dos fósseis responde à segunda pergunta (o “Quando chegaram à ilha”), já a sistemática e taxonomia efetuada por reputados especialistas responde à terceira questão, a qual lida com os fenómenos de especiação. Falta a resposta à primeira questão, na qual se abordam as origens dos colonizadores. É aqui que entra, mais uma vez, a Biogeografia Insular e o estudo das relações biogeográficas dos organismos terrestres, bem como dos marinhos. Para responder cabalmente a esta importante questão, é necessário conhecer em detalhe as listas de espécies, validadas por especialistas, dos organismos selecionados (por exemplo, moluscos marinhos gastrópodes, equinodermes, algas) em cada local a comparar. É aqui que adquirem relevo as coleções curadas em Museus, as quais, após compiladas, acabam por preencher os requisitos para serem incluídas nos processos de Big Data.
Por definição, Big Data é uma área do conhecimento que se dedica ao estudo de um grande volume de dados. Nos últimos anos, temos assistido a uma cada vez maior utilização de Big Data pelos biólogos, seja através da construção de grandes bases de dados taxonómicas para organismos recentes (por exemplo: WoRMS — World Register of Marine Species) ou passados (isto é, de fósseis; ex: PaleoBiology Database), seja, por exemplo, através da construção de árvores filogenéticas utilizando genomas completos de vários organismos, o que requer capacidade computacional cada vez mais substancial. Consequência disto, é a procura crescente por parte do mercado científico e empresarial de cientistas (programadores, algoritmistas, engenheiros de sistemas) capazes de darem respostas aos problemas com que os investigadores se deparam na análise de volumes de dados cada vez maiores.
A par destes desenvolvimentos recentes nesta nova área do conhecimento que é o Big Data, assiste-se no nosso país a um recrudescimento do investimento em Museus, com o correspondente reconhecimento da importância e valorização da curadoria das coleções biológicas, paleontológicas e geológicas que são mantidas a cargo destas instituições. Dou aqui dois exemplos de sucesso: o Museu de História Natural e da Ciência da Universidade do Porto, instituído formalmente em 2015 e em funcionamento desde 2019 e a Casa dos Fósseis, inaugurada em 2016 em Vila do Porto, onde é divulgado ao público o extraordinário e internacionalmente relevante património paleontológico existente na ilha de Santa Maria, no arquipélago dos Açores (FIGURA 1).
Estas coleções são fundamentais, não só como repositório da biodiversidade atual e passada — os grandes Museus são muito visitados por cientistas porque possuem enormes coleções e muito exemplares-tipo (os quais correspondem aos indivíduos que serviram para a descrição formal de uma dada espécie) — mas também pelo relevante papel educativo que desempenham ao promoverem a Ciência e o conhecimento.

Vamos exemplificar tudo o que atrás foi dito, através de um caso de estudo. A Macaronésia é uma ampla área geográfica localizada no Atlântico norte, onde estão cinco arquipélagos: Açores, Madeira, Selvagens, Canárias e Cabo Verde. No total, nesta região, existem 31 ilhas de maior dimensão e muitos ilhéus e montes submarinos (FIGURA 2).
A palavra “Macaronésia” foi cunhada pela primeira vez pelo botânico britânico Philip Barker-Webb (ca. 1845) para abranger os arquipélagos da Madeira, Selvagens e Canárias. Mais tarde, Engler, em 1879[16], incluiu os Açores na região da Macaronésia e Dansereau, em 1961[17], alargou ainda mais o conceito, ao incluir as ilhas de Cabo Verde. Alguns autores consideram que outras regiões apresentam também um número significativo de taxa comuns com as ilhas da Macaronésia, nomeadamente algumas áreas da Península Ibérica e algumas zonas costeiras do noroeste africano adjacente. Embora o termo “Macaronésia” tenha sido usado com diferentes significados, a inclusão de Cabo Verde é um assunto particularmente controverso, em especial para os especialistas no meio marinho[18].
Vários estudos baseados em peixes marinhos costeiros e gastrópodes de Cabo Verde concluíram que a estrutura da comunidade e as relações biogeográficas do biota marinho deste arquipélago diferem significativamente dos outros arquipélagos da Macaronésia, mas, até recentemente, esta questão não tinha sido abordada de forma quantitativa, estruturada e através de um estudo multifilo. Enquadrado pelas premissas do modelo dinâmico Sea-Level Sensitive (SLS) da biogeografia insular marinha[19] e fundamentado nos processos e padrões evolutivos biogeográficos insulares, foi recentemente publicado o estudo mais abrangente, até à data, acerca da biogeografia marinha macaronésica[20].
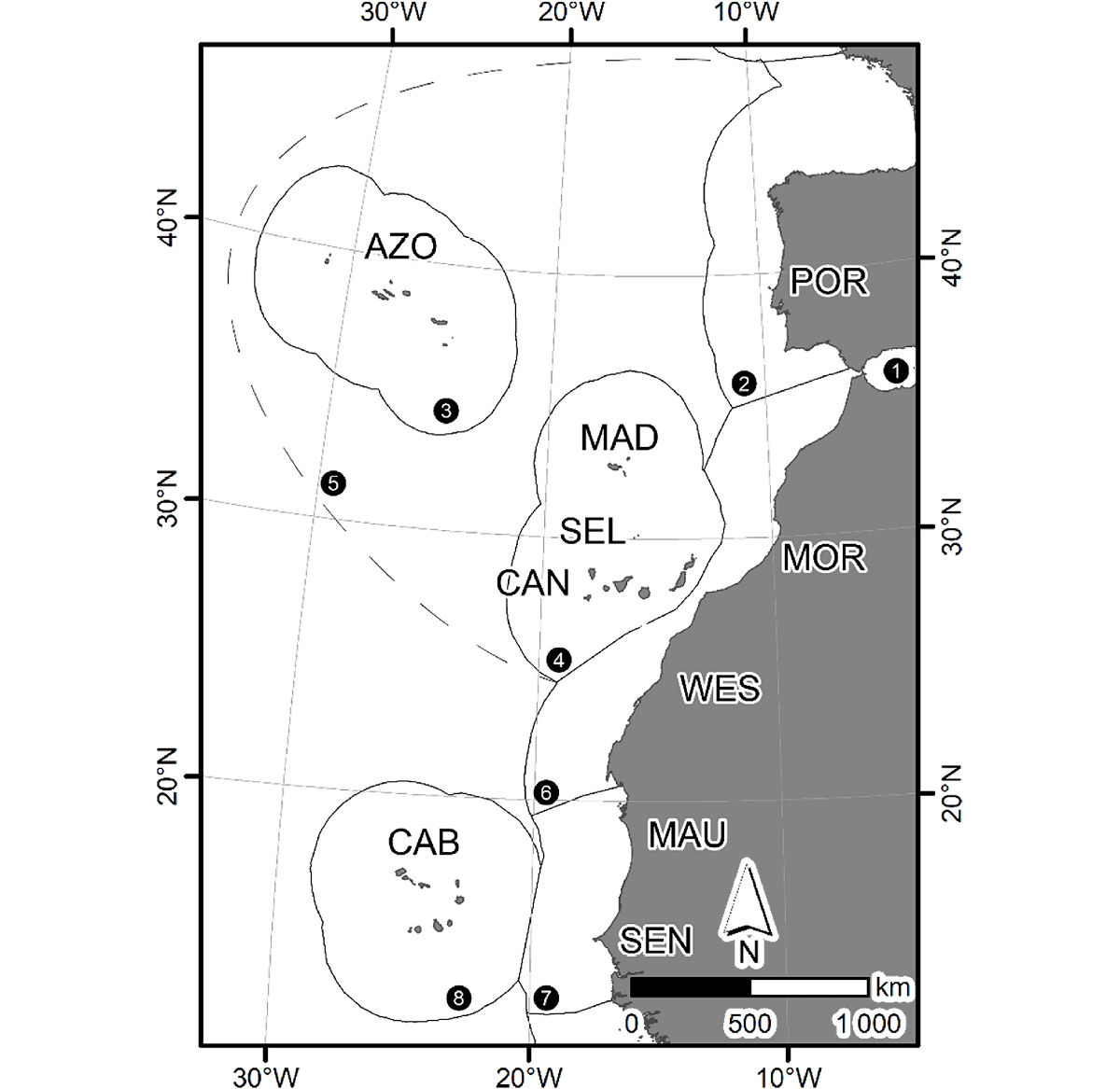
Coordenado por um dos autores (SPA), este trabalho permitiu efetivar, do ponto de vista marinho, a primeira análise comparativa multifilo para reavaliar este debate e, assim, testar se a Macaronésia era (ou não) uma unidade biogeográfica coerente. Para tal, efetuou- se uma compilação exaustiva de listas de espécies, avalizadas por especialistas, dos seis grupos nativos marinhos macaronésicos mais estudados: peixes costeiros, equinodermes, moluscos gastrópodes, crustáceos decápodos braquiúros, anelídeos poliquetas e macroalgas. Foram ainda incluídas outras regiões próximas (por exemplo, Portugal continental, o Mediterrâneo, ilhas Britânicas, entre outras), as quais serviram para melhor elucidar as relações biogeográficas entre os vários arquipélagos que formam a Macaronésia, e entre a Macaronésia e as outras regiões selecionadas. No total, foram incluídas 7 492 espécies marinhas, sendo que 3 737 destas ocorrem nos arquipélagos da Macaronésia. A principal conclusão deste trabalho levou à criação de uma nova unidade biogeográfica — a ecorregião da Webbnesia — assim homenageando Philip Barker-Webb e, pela primeira vez baseada em dados quantitativos sonantes, permitiu individualizar e separar os arquipélagos dos Açores e de Cabo Verde dos restantes (FIGURA 3).
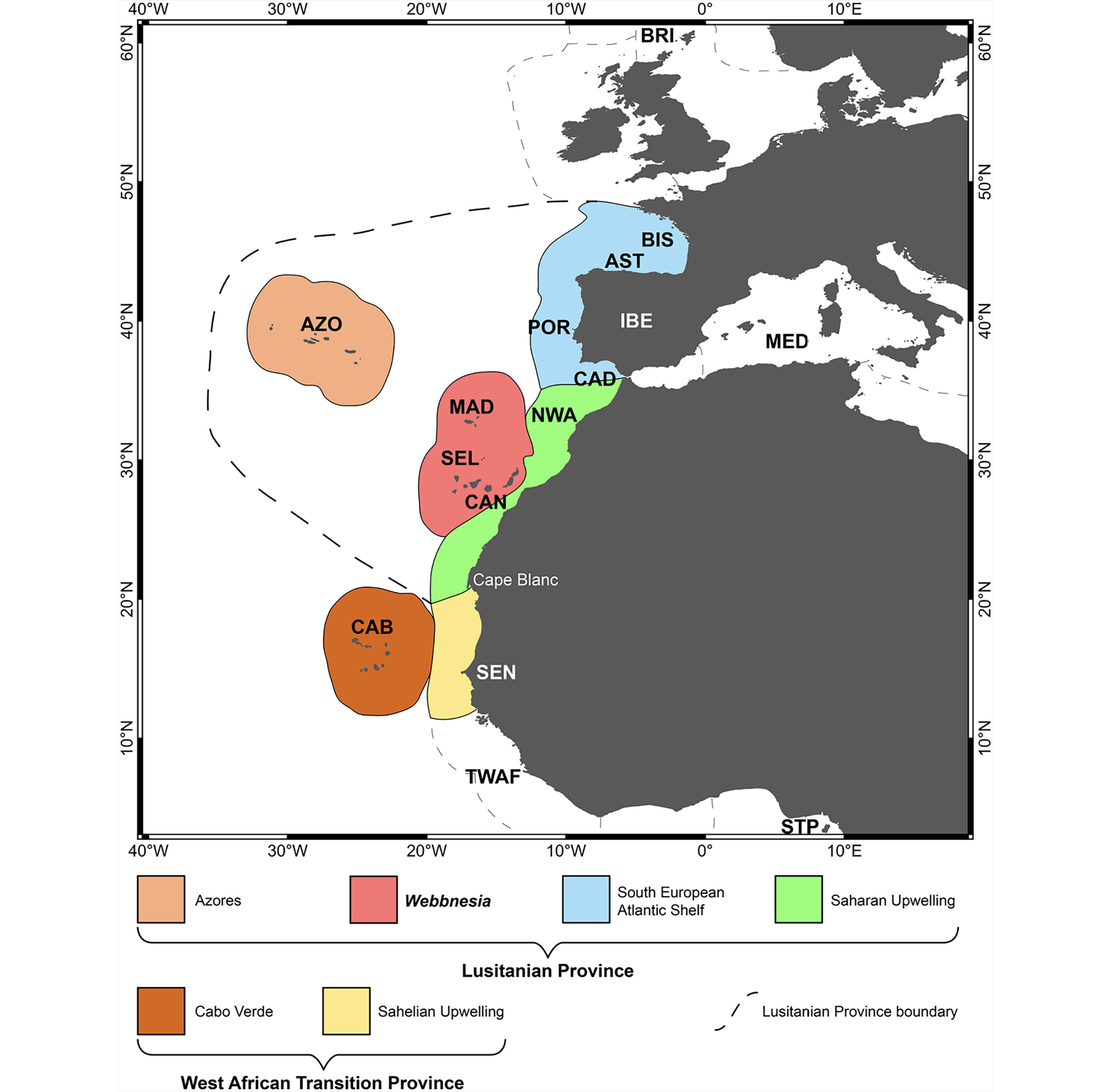
Assim, Cabo Verde deverá ser considerado como uma subprovíncia biogeográfica isolada, os Açores como uma ecorregião e Madeira, Selvagens e Canárias formam uma nova ecorregião, denominada Webbnesia. É este o poder do uso de Big Data!
Referências
- ↑ PATIÑO, J. et al., A roadmap for island biology: 50 fundamental questions after 50 years of The Theory of Island Biogeography, Journal of Biogeography, 44, 963–983. 2017. DOI: 10.1111/jbi.12986.
- ↑ RAMALHO, R. et al., Coastal evolution on volcanic oceanic islands: a complex interplay between volcanism, erosion, sedimentation and biogenic production, Earth Science Reviews, 127, 140-170. 2013. DOI: 10.1016/j.earscirev.2013.10.007.
- ↑ ÁVILA, S. P. et al., Towards a “Sea-Level Sensitive Marine Island Biogeography” model: the impact of glacio-eustatic oscillations in global marine island biogeographic patterns, Biological Reviews, 94, 3, 1116-1142. 2019. DOI: 10.1111/ brv.12492.
- ↑ WALLACE, A. R., Island Life: Or, the Phenomena and Causes of Insular Faunas and Floras, Including a Revision and Attempted Solution of the Problem of Geological Climates, Macmillan & Co, London. 1880.
- ↑ MACARTHUR, R. H. & WILSON, E. O., The Theory of Island Biogeography, Princeton University Press, Princeton. 1967.
- ↑ WHITTAKER, R. J. & FERNÁNDEZ–PALACIOS, J. M, Island Biogeography: Ecology, Evolution and Conservation, 2nd Edition, Oxford University Press, Oxford. 2007.
- ↑ FERNÁNDEZ-PALACIOS, J. M. et al., Towards a glacial–sensitive model of Island biogeography, Global Ecology and Biogeography, 25, 817–830. 2016.
- ↑ BORREGAARD, M. K. et al., The general dynamic model: towards a unified theory of Island biogeography?, Global Ecology and Biogeography, 25, 805–816. 2016.
- ↑ BORREGAARD, M. K. et al., Oceanic Island biogeography through the lens of the general dynamic model: assessment and prospect, Biological Reviews, 92, 830–853. 2017.
- ↑ WHITTAKER, R. J. et al., Island biogeography: taking the long view of nature’s laboratories, Science, 357: eaam8326. 2017.
- ↑ ÁVILA, S. P. et al., Towards a “Sea-Level Sensitive Marine Island Biogeography” model: the impact of glacio-eustatic oscillations in global marine island biogeographic patterns, Biological Reviews, 94, 3, 1116-1142. 2019. DOI: 10.1111/ brv.12492.
- ↑ HACHICH, N. F. et al., Island biogeography: patterns of marine shallow-water organisms in the Atlantic Ocean, Journal of Biogeography, 42, 1871–1882. 2015.
- ↑ ÁVILA, S. P. et al., Comment on ‘‘Island biogeography: patterns of marine shallow-water organisms’’ by Hachich et al., journal of biogeography (2015), Journal of Biogeography, 43, 2515–2516. 2016.
- ↑ ÁVILA, S. P. et al., Global change impacts on large-scale biogeographic patterns of marine organisms on Atlantic oceanic islands, Marine Pollution Bulletin, 126, 101–112. 2018.
- ↑ PINHEIRO, H. et al., Island biogeography of marine organisms, Nature, 549, 7670, 82–85. 2017.
- ↑ ENGLER, A., XXX, Die extra-tropischen Gebiete der nördlischen Hemisphäre, Engelmann, Leipzig, Germany. 1879.
- ↑ ANSEREAU, P., Études macaronésiennes I: géographie des Cryptogames vasculaires, Agronomica Lusitanica, 23, 151–181. 1961.
- ↑ WIRTZ, P. et al., The coastal fishes of the Cape Verde Islands - New records and an annotated check-list: (Pisces), Spixiana, 36, 113-142. 2013.
- ↑ ÁVILA, S. P. et al., Towards a “Sea-Level Sensitive Marine Island Biogeography” model: the impact of glacio-eustatic oscillations in global marine island biogeographic patterns, Biological Reviews, 94, 3, 1116-1142. 2019. DOI: 10.1111/ brv.12492.
- ↑ FREITAS, R. M. et al., Restructuring of the “Macaronesia” biogeographic unit: a marine multi-taxon biogeographical approach, Scientific Reports, 9, 15792. 2019. DOI: 10.1038/s41598-019-51786-6.
- ↑ SPALDING, M. D. et al., Marine ecoregions of the world: a bioregionalization of coastal and shelf areas, Bioscience, 57, 7, 573e583. 2007.
- ↑ FREITAS, R. M. et al., Restructuring of the “Macaronesia” biogeographic unit: a marine multi-taxon biogeographical approach, Scientific Reports, 9, 15792. 2019. DOI: 10.1038/s41598-019-51786-6.
Criada em 23 de Novembro de 2022
Revista em 23 de Novembro de 2022
Aceite pelo editor em 20 de Dezembro de 2022